





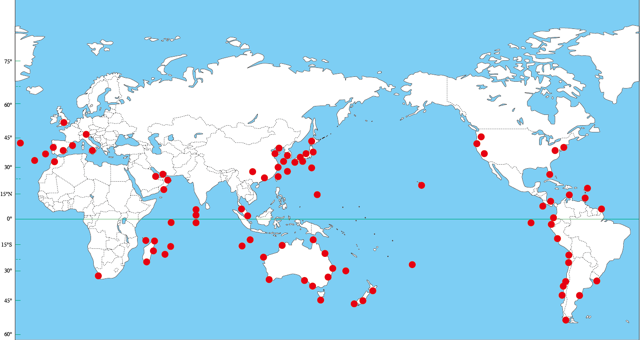
 |
2018.1.19
No.26�@�u���R���̃��[�c��K�˂āv
���R�������̗��j
���R�������Ƃ́A�{���͍��R�т݂̂őS���C�t�T�C�N���������������w���ׂ��ł��낤���A���ނł͊���I�Ƀ~���}�V���`���E�A�N���}�c�}�L�`���E�A�~���}
�����L�`���E�A�I�I�C�`�����W�A�R�q�I�h�V�A�x�j�q�J�Q�A�N���}�x�j�q�J�Q�A�^�J�l�q�J�Q�A�^�J�l�L�}�_���Z�Z����9��ނ����R���ƌĂ�ł���B���a����
���r�������ޓ��D��̉ZEPHYRUS�n�����̊�������������R���ł������i�}1�j�悤�ɁA�����������璱�ވ��D�Ƃ̔M��������������Ă����B
���R���̔����́A���{�A���v�X�o�R�j���t�����Ƌ@����ɂ��A�����N�Ԍ㔼�ɏW�����Ă���B�����̃A���s�j�X�g�̑����͔����w�ɂ����w���[���A���{�R�x���
�������o�[�V�l�̒��S�l�܂ł����{�������u��̒��S�I�l���ł������B����43�N(1910�N)�A���̑咬�s������㗧�R�A���R�A��j����r����
�N���}�c�}�L�`���E�ƃ^�J�l�q�J�Q�����Ŕ����������������Y�́u���R����������v�i�c���s�j�F���R�������ɍĘ^,
1959�j�́A�K�C�h�����Ȃ��R�ł𑖔j���郍�}���ɖ������T���I�o�R������A�M�������Ƌ��ɍ��ɓ`���閼���ł���B�~���}�V���`���E�i1901
�N�j�A�N���}�x�j�q�J�Q�i1905�N�j�A�^�J�l�L�}�_���Z�Z���i1915�N�j�Ȃǂ����O�サ�Ĕ�������A���R���Ƃ����ď͍̂���鑠�i�R�x�n����,
1906�j���ŏ��ł���Ƃ����B�x�j�q�J�Q�̔����͑����āA��ԎR�Y�Ɋ�Â���1877�N�ɋL�ڂ���Ă���B
���̌�̌����ō��R�������ƌĂׂ�̂̓^�J�l�q�J�Q�������ł��邱�Ƃ������������A���܂ł��ꊇ���č��R���ƌĂѐe���܂�Ă���B�C�O�ł̓��[���V�A��
���̒��ܓx�n�т̍��W���n��A���ܓx�n�тł͕��n�ɍL�����������ł���B���R�A���Ɠ����悤�ɖk���N���̐����ŁA���{���嗤�Ɨ������ł������X�͎�
��ɓ��{�֓n���Ă����ƍl�����Ă���B�����������̕X�͎���Ȃ̂��A�܂��T�n�����Ȃǖk���o�R���A���N�����o�R���ȂǏ�������N����Ă����B
���R���Ƃ̏o�
���R������ʂɈӎ��������������͎G���u�V�����v1956�N7�����́u�M�B���W���v�ŁA�\���ɂ̓n�C�}�c�Ɏ~�܂�^�J�l�q�J�Q�̎ʐ^���ڂ��Ă����B���m�N
���Ȃ��狭���S�䂩�����̂�����A����ɕ������R���̎p��z������Ƌ����������B���ۂɏo������̂͂��ꂩ�琔�N��̑�w����ŁA�����R�x�n�����̏W
���ĉ��A�����̍��R���ɐڂ��邱�Ƃ��ł����B
���x�o�ϐ��������s�[�N���}�������ɂ́A���~�x�݂��Љ�ŔF�m������Ƃ肪���܂ꂽ�B�x�ɂ𗘗p���ĉƑ��Ƌ��ɒ����R�x�Ⓦ�k�n���̎R�X��K���ƁA��
�̎����ɂ͍g�F�̔��䂪�N�₩�ȃx�j�q�J�Q�i�}2�j���}���Ă��ꂽ�B���F�⎇�̍��R�A���Ɍe�����̎p�́A���́u�M�B���W���v�̊�����h���点�Ă��ꂽ�B
�e�n�̕W�{���W�܂��Ă���ƁA������ł�����Ȃǂɒn���I�ȕψق����邱�ƂɋC�����B��������Ǝ��ɂ͕ψقݏo����������m�肽���Ȃ�A�Ȃ����̂�
���Ȓn���I���z�����Ă���̂��낤�Ǝ��X�ƍD��S���N���Ă���B�����w�ւ̑z���ł���B�x�j�q�J�Q�͒n���I�ψق�̕ψقɕx��ł��āA1950�N��ɂ�
�����悤�Ɉ��햼�������A�����K���L���Ȉ��햼�͏\���ɂ��̂ڂ�B���̕ψقɖ�����ꂽ�����҂∤�D�Ƃ������A�Â�����ψق̌n���ƒn���I���z�Ɋ�
����l�@���s���Ă����B�k�C���Y�ł́��ɔ��������邪�{�B�Y�ɂ͂قƂ�ǔF�߂�ꂸ�A�܂����O���̍g��̏o���ɂ������ȍ��������āA�ʎ����������l
������B�܂����k�n���̎�v�ȍ��R�ɕ��z����ɂ�������炸�A���b�c�R�Ȃǂ̍��R��i����X���ɕ��z���Ȃ��̂��s�v�c�ł���B
����𐔒l�������v�����ɂ���Ă��̓�������͂���A�e�n�Y�̌Q�̌n���W���\�z�ł���B���̌n���W���番�z�g��̕��������T��Ȃ����낤���B�x��
���Ƃɔ��䑪��Ɠ��v�������s���A�Ƃ肠�������k�n���e�n�Y�̌n���W�𑽕ϗʉ�͖@�ł܂Ƃ߂��i1996�j�B����Ɋe�n�Y�̓��v������ςݏd�˂Ă���
�ƁA�����R�n�ł��W���̍����Ƃ���ɕ��z����̌Q�́A�T�C�Y���������A�x�j�䂪�k�����A���䂪�������Ȃ�Ƃ����@�����̂��邱�ƂɋC���t�����B���v
��������Ȃǂ̌`���ɏ��Ȃ��炸�e�����Ă���悤���B����ł͌`���I�ȓ�����P���ɏ������Čn���W���]�X���邱�Ƃ͂ł����A���ϗʉ�͂̋Z�@�����낢��
�g�ݍ��킹�đ����I�ɕ]������K�v������B����ƌ`�����Ɋ�Â��n���W���\�z���邱�Ƃ��ł����̂�2013�N�̂��Ƃł���B��ɏq�ׂ镪�q�n���I�Ȏ�
�@�ɂ��n���n���I���ʂƔ�r����ƁA��M�z�n��̈ꕔ�������Η��҂��悭��v���邱�Ƃ������ꂽ�B����ψق͈ꌩ�����_���ɐ����Ă���悤�Ɍ����邪�A
�K�Ȑ����������{�����Ƃʼnߋ��ɐ������n���Ɉˑ�����ψٌX���𒊏o�ł����̂ł���B�ŏ��ɓ��v�����̕\�����̂�1984�N�ł��邩��30�N��
�Ό����o���Ă����B
�{�B�̃x�j�q�J�Q�̐����n�̂قƂ�ǂ��ׂĂ�K��āA���̐��������ώ@���Ă���Ɩʔ������Ƃ��������Ă����B���k�n���Ńx�j�q�J�Q�̐�������R�X�́A�u
�i�Ȃǂ̍L�t���ёт̏㕔�ɒʏ�͑��݂���j�t���ёт������Ă���A����ɋU���R���ƌĂ�鑐�����L���邱�ƂŁA�W�����Ⴂ�ɂ��ւ�炸�C�l�Ȃ�X�Q
�Ȃ�H���Ƃ���x�j�q�J�Q�̐������\�ɂ��Ă���B�ĐΘA���a��A��̂悤�ɋU���R�������̉����ƁA�R�̒���Ƃ̍���100-200m�����Ȃ��R�n����
��B
2000�N�̍��܂ł́A�ŏI�X���ȍ~�̍ł����g�Ȏ����i�q�v�V�T�[�}���j�ɂ͌��݂Ɣ�r���Đj�t���ёт���400m�㏸���Ă����Ƃ����̂��X�ѐ��Ԋw�E��
����ł��������A���ёт�400m���㏸������A�������Ƌ��Ƀx�j�q�J�Q�͐�ł����͂��ł���B���k�n���̃x�j�q�J�Q�̕��z�ɂ͓��Ă͂܂�Ȃ�����
��2001�N�ɘ_���Ŏw�E�����B���̌�A�U���R�������̂���R�X�ɂ́A�܂��j�t���т��i�o���Ă��Ȃ��Ƃ���V���Ȋw�����L�͂ƂȂ�A���k�n���̃x�j�q�J�Q
�̕��z�Ɛ��������悤�ɂȂ����B�H�A�������͐H�����܂ސA���������łȂ��Òn����ËC������L���鐶���Ƃ̋����Ő��������`�����Ă���̂ŁA�t�B�[��
�h�E�f�[�^����ɑ����I�ɍl���邱�Ƃ̏d�v�������߂Ċ���������ꂽ�B
�x�j�q�J�Q�̕��z����͓�A���v�X�̓�[�n��ɂȂ邪�A���̕t�߂Ɉʒu����喳�ԎR�Ȃǂ͎R��S�̂������R���j�t���тŕ����Ă���B��ʓI�ɂ̓x�j�q�J
�Q�̐����ł��Ȃ����ł��邪�A���̋������n��A�}�s�ȕ���n�̑J�ړr��̑��t�����������x���Ă���B���̂悤�Ȑ������́A�k�����̃x�j�q�J�Q����
�g���ɐ����n���k�����Ȃ��琶���c�������t���W�A�̊���A�z�����Ă����B
���q�n���n���w�Ƃ̏o�
1970�N��ɂ͓��{�̐��������Ɗ֘A�����Ē��̕��z�p�^�[����_���邱�Ƃ����s�����B�����Ȓ��ތ����Ƃ̔��������́u�Ƒ嗤��K���ɕ��f�E�ڑ�������A�ǂ�ȕ��z�p�^�[���ł�����v�ƈ��Ղȕ��z�`���_�̗��s�Ɍx����炳�ꂽ�B
1995�N��������V�ȎO����ɂ��I�T���V�̕��q�n����͂����\�����悤�ɂȂ����B���̎c��Ȃ������̐i���̗��j���A�������DNA����H�邱�Ƃ�
�ł���A�Ƃ����̂͐��ɖ��͓I�ŁA���̕���ł��������Ă݂����Ƃ̎v���ɋ��ꂽ�B���̍��C���^�[�l�b�g�̏��T�C�g�ŁA�����O�O��w�ɂ���ꂽ�F����
�^�ꎁ�i���M�B��w��w�������j�����q�n����͂̃T���v���Ƃ��ăx�j�q�J�Q�����߂Ă���ꂽ�̂ŁA�e�n�̃T���v���������肵���B�܂�DNA��͋Z�p������
���m�l�̊����K���Ƌ����Ńx�j�q�J�Q�̌������n�߂��Ƃ���V��ׁE�ؕ鐉���������łɒ��肳��Ă��鎖��m��A�����Ő��ʂ̈�[��2000�N�ɓ��{����
�w��ɔ��\�����B�܂�2003�N�Ɋ�b�����w�������ŊJ�Â��ꂽ�u�����̐i�����l���ƕ��q�n���v������ł́A�F�������������쌧�̍��R���ɂ��Ĕ��\��
��A���k����@����Ă��͍̂K���ł������B
���̂悤�Ȃ�������A2004�N�Ɏv�����ĐM�B��w��w�@���m�ے��i���w���ɓ����v�������j�ɓ��w���A���{�s�ɃA�p�[�g����đ�w�ʂ����n�܂����B
�e�[�}�̓x�j�q�J�Q�̓��{�ɂ����镪�z�ϑJ�j�̉𖾂ł���B���{�Y�̃x�j�q�J�Q�̕��z�`���j�͊�����ƒ���ς������̂ʼn𖾂ɂ͂���قǎ��Ԃ�v��
�Ȃ��Ǝv���Ă����̂ŁA�v�攭�\�ł͐��E�̃x�j�q�J�Q���̕��q�n�������荞�B
���{�s�͓��Ƀs���~�b�h�^�̏�O�x�����Ƃ���k�A���v�X�̍��A�Ȃ�A���ɂ͔������̂Ȃ��炩�ȕ������B�l�G�܁X�Ɍ�����R�X�̎p�͌��O���邱
�Ƃ��Ȃ��B��O�A��͑����̍��R���̐��݂��œ~�̊Ԃ͊��Ⴗ��̂ɑ��āA�������͍��R�������z�����~�ł��قƂ�ǐႪ�ς���Ȃ��B���̑ΏƓI�ȘȂ܂���
�݂Ă���ƁA���R���̕��z�Ɛϐ�ɉ������ւ�����Ǝv���ĂȂ�Ȃ��B���R���̐��݉Ƃ܂œ��A��\�Ƃ����b�܂ꂽ���ŁA�����̖����Ⴂ�@���Ƌ��ɐi��
�����w���w�Ԃ��Ƃ͊y���������B�V���̐i���]�Ŗ��T�̃[�~�����Ȃ��̂͑�ςł��������A�Ⴂ�����̉@���͂��ׂĐ搶���œۂݍ��݂̈����V�l�̖ʓ|����
�����Ă��ꂽ���Ƃ͊��ӂɑς��Ȃ��B3�N�ԂɊw�i�������w�̒m���͂�������A�������n���w��ËC��w�ɂ��āA���ꂼ����̐搶����ŐV�̊w
�������f���ł��������b�g�͌���Ȃ��傫���B��ɐi������W�������̍ŐV�̒m���͂Ȃ��Ȃ��Ɗw�ł͓����Ȃ����̂ŁA��w�i�@�j�̑��݈Ӌ`�͂܂��ɂ�����
���w�ۓI�m���̓�����Ƃ���ɂ���Ǝv���B
�x�j�q�J�Q��DNA��͂ɂ��ẮA�F���������̋��ŊǗ�����Ă��钷�쌧�Y�̃T���v����A�ȑO������W���Ă����W�{���g�p�ł������A�ꕔ�̒n��Y�͍ēx
�̂蒼���˂Ȃ�Ȃ������B�Ȃɂ���N���A�Ă̈ꃖ���サ�����������A���������R�т܂œo��ɂ̗͑͂��K�v�ł��邵�A�Ȃɂ��V�C�Ɍb�܂�Ȃ���Β���
��Ȃ��B�܂����ʕی�n��ł͋����K�v�ł���B�Z������̂��߈ɓ������ƉF�����������͂��߁A���{�ނ��̉�̕��X�ɂ�����Ȃ����͂����B
���ׂĂ݂�ƃx�j�q�J�Q�͖�27���N�O�ɁA�{�B�ł͓�k2�̌n���ɕ��A���̌���̕X�̓T�C�N���ɑΉ����ĕ��z�̊g��i������j�E�k���i���g���j��
�J��Ԃ������Ƃ����������B�{�B�Y�͓�A���v�X�Ɩk�A���v�X�̃��t���W�A�Ő��B�I�Ɋu�����ꂽ���Ƃ�2�n���ɕ��A���̌㕪�z���g�傷��ߒ��œ��k�n��
�Y�Ɠ�A���v�X�Y���k���n���ɁA���R���܂ނ��̑��̒����R�x�Y������n���ɕ����̂ł���B�܂���k2�n���́A��z�R�n�̒J��A��ō������Ă��鎖����
���������A���̒T���s�͖{��No.7(2007)�ɏ������B
���R���̃��[�c�����߂�
�x�j�q�J�Q���Ђƒi������ƁA���̑��̍��R���ɂ��ڂ������Ă����B�{�B�̍��R���̃��[�c��T�邤���ŃL�[�ƂȂ�n��́A���������{�̑Ί݂Ɉʒu����
�A���[���n��ƒ��N�����k�����ӂł���B�������A�e�킲�Ƃɑ嗤�ł̈�`�I�ψق̃p�^�[����c�����邱�Ƃ́A�ւ̓n����������ł̕��z�ϑJ�̓���
�𗝉������ŏd�v�ł���B
��N�ސE���2000�N����ɁA���N���[���V�A�Ɩk�ĂŃT���v�����W�ɓw�߁A�����^�|���ɂ͑����̒����s�œ��s�����Ă��������A��ɂ͂�������q����
���B�������l�ōL���嗤��ԗ��I�ɒ�������킯�ɂ����Ȃ��̂ŁA�����̕����炲���͂������Ƃ͂����܂ł��Ȃ����A�����̃R���N�^�[�C���͗}������
���B�������A������n�������s�����Ƃ̏d�v���́A�t�B�[���h�E���[�N���|�Ƃ��Ă�����F����ɂ͂悭��������Ǝv���B�Ⴆ�A�~���}�����L�`���E��
�͌����Ȉ�`�q�^��2�n�������āA�e�n�ō������Ă��邱�Ƃ������������A���N�[�e�B�A�̃E�X�`�E�l���ł̓s���|�C���g�œ���̏ꏊ�ō̏W�����T���v������
2�n�������������B�����u�����v�Ƃ����\���ł��A����̏W�����T���v���ƁA�����T���v���ł͏d�݂��قȂ�̂ł���B
���{�ł͍��R���ł��嗤�ł͕��ʎ킾�Ƃ悭�����邪�A�L�敪�z�킪���Ȃ炸�������ʎ�Ƃ͌���Ȃ��B�I�I�C�`�����W�̓A���[���n��ł͖L�Y���邪�i�}
3�j�A���[���b�p�̃I�I�C�`�����W�Ȃǂ͂��̕��ނłȂ��Ȃ���ɓ���Ȃ��B�u���K���A�ł̒����ŁA�Ԃňړ����ɋx�e���Ă�����A���܂��ܐ����c��̎��ɑ�
�������Ȃǂ́A�܂������F��Ƃ��������悤���Ȃ��B���[���b�p�Y�ŗB��̃T���v���́A���̂Ƃ�����ꂽ���̂��B
�{�B�Y�̍��R�����A�T�n�����E�k�C���o�R�Ŗk���ɓ��B�������̂��A���N�����o�R�œ���ɓn���������̂��B�k���n�̐����Ƃ������ƂłȂ�ƂȂ��k����
�l���Ă��܂����A�ʂ̎��_���K�v�ł��邱�Ƃ��w�E�����͍̂���ǕF���i���R�̒������A���R�Ȋw�Ɣ�����46(2),
1979�j���B���͖{�B�̍��R���̑������k�C���ɎY���Ȃ����Ƃ�A�A���[���n��Y�Ƃ͌`�ԓI�ɔ��ɕς�����킪���邱�ƂȂǂ���A�ꕔ�̎�͓���œn
�������̂ł͂Ȃ����ƒ�N�����B
�T�n�����Y�̃T���v���ɂ��ẮA���炭�T�n�����̒�����T������Ă��钩�����ꎁ��葽���̃T���v������Ă��������A2010�N���ɂ͂��ׂĂ̎��
��͂��I���Ă���A��������A���[���n��Y�ƈ�`�q�I�ɂ͕ψق������Ȃ����Ƃ��c���ł��Ă����B����͂ǂ��������Ƃ��낤�B�T�n��������k�C���֓n����
���Ȃ������̂��A���邢�͓n���������k�C���ł͐�ł������B�܂��͒��N�����o�R�Ȃ̂��낤���B
���낢��l�������点�Ă������A�]�@��2012�N�ɓˑR�K�ꂽ�B�T�n�����ŃI�I�C�`�����W���̏W���ꂽ�̂��B���N�ɂ͎����܂ޕ������������ēy����ł�
�邱�Ƃ��m��I�ɂȂ�A�T���v�������Ē������B���ʂ͔��т̍L���`�Ԃ��琄�肳�ꂽ�ʂ�A�A���[���n��Y�Ɠ���̃n�v���^�C�v�Ŗk�C���Y�Ƃ͈قȂ���
�����B�k�C���e�n�̃I�I�C�`�����W�̃n�v���^�C�v�͋ψ�ŁA�A���[���n��Ɠ���̃n�v���^�C�v�͌��o���Ȃ��̂ŁA�T�n�����܂ŐN�������̌Q�͖k�C���ւ�
�n���ł��Ȃ������̂ł͂Ȃ����낤���B
�{������̑�13���W��i���{�s,
2016�j�̌��J�V���|�W�E���ŁA����܂ł̒m���\�����B���ɘ_���Ƃ��Ĕ��\�����x�j�q�J�Q�ƃN���}�x�j�q�J�Q�́A���R���̒��ł��嗤�Y�Ɠ��{�Y
�̈�`�I�����͌Â��A���\���N����100���N���炢�O�ɓ��{�֓n�������ƍl������̂ɑ��āA���̑��̍��R���͂����ƐV��������ɓn�������Ɛ��肳
���B�n�����[�g�����҂ł͂��Ȃ葊�Ⴊ����悤���B���̗v�������낢��Ƒz�����邱�Ƃ́A���߂č��R���ɏo��������N����̎v���o�ɂȂ����Ċy�����B
�]���O���́u���R�����l�v�i�j���[�E�G���g�����W�X�g2(1/2), 1952�j�̒��ŁA�u���݂̕��z�͕��G�ł��邪�A���̐����ɂ͋��R�ɂ����̂������ƍl������v�Ə����Ă����邪�A�^�Ɋܒ~�ɕx��ł���Ƃ��킴��Ȃ��B
 �@�@�@ �@�@�@
�}�P�[�t�B���X�n�����̑}�}�i1929�j
�ォ��~���}�V���`���E�A�N���}�c�}�L�`���E�A�^�J�l�q�J�Q

�}�Q�@������̃x�j�q�J�Q
��z�R�n�A���@�R�ɂāi�r�c�^��Y���B�e�j

�}3�A�ʂɏW�܂�I�I�C�`�����W
���V�A�A�R���\�����X�N�E�i�E�A���[���x�O�ɂ�
|
���J �M��
�@
�Ó쐶���n���w������
(�J�i�_�k�����B�A�k�Ɍ��̃}�b�P���W�[�E�f���^��w�Ɂj
|
2018.1.19
No.25�@�u�����̏W���琶���n���Ɓg��h�̖��ցv
���Z�ŗ��ȁi�����j�S���̋��t�����Ă�������1994�N�ɒ�N�ސE���A���݂͋��N�i2016�N�j�É��s�ɒa�����������́g�ӂ��̂��ɒn�����j�~���[�W�A���h�ō����ށi���ށj�̕W�{�E�����̐����𑱂��Ă��܂��i�ʐ^�P�j�B
�@�c�����ɉ߂������R�`���E�O�d���Œ��̂���n�߁A���̌�A�É��s��1950�N���Z�ɐi�w���A���������Ƃ��Ē��ނ̍̏W�E�ώ@�ɖv�����܂����i�ʐ^�Q�j�B
�@���Z����ɑ��Z�̐������Ƃ��𗬂��A�����ŏW�߂����������ƂɁA�u�É������g�����C�ݒn�т����A���v�X��x�m�R�̍���n�܂ŋC��̕ω��ɕx�݁A�����
�قڕx�m������E�Ƃ��āA���ɔ�ΎR�n��A���ɉΎR�n�悪���邱�ƁA�����Ē��ޕ��z�����̕ω��ɕx���R�������ɔ��f���Ă��邱�Ɓv��m��܂����B
�@���ꂪ���̐����n���w�ւ̖ڂ��߂������̂ł��B���̒��͂Ȃ������ɐ��ނ̂��A�Ƃ������Ƃ����̃e�[�}�ƂȂ�܂��B
�@���͒n���̐É���w�����w���ɓ��w���A�����w���U���܂����B�����ĂЂ�ς�ɖ�O�ɏo�����A���Z����ɒ��z�������ނ̐����n���w��[�߂�w�͂𑱂��܂����B
�@��w�ł́A�A�������w�̖؉��O�Y�搶����u�ЂƂ̎����⌻�ۂɋC�Â�����A�Ȃ������Ȃ̂��������ł��悢���痝�������Ă݂Ȃ����A�����Ԉ���Ă���
��V�����������l����Ηǂ��̂ł͂Ȃ����v�A�n���w�̎L���P�F�搶����u���R�Ȋw���w�Ԏ҂͑傢�Ɍ|�p�ɐe���݊���{���Ȃ����B�������琶�܂�钼��
���V���������ɂȂ���̂��v�Ƃ������ƂȂǂ��w�т܂����B
�@����w����w���̓����������Ԋw�E��t�ΐ��搶�̌������ł́A��B��w�̔��� ���搶�́A���ޕ��z�ɂ�����g�����x�ߌn�h�̏d�v���ɂ��ẮA���̗L���Ș_���ɏo��A���̒���Ș_�����m�[�g�ɏ����ʂ��A������ËL����قǂ���Ԃ��ēǂ��̂ł����B
�@���͓s���ɂ��Z���̔����Y�搶��ьc�搶�������Â˂��A���낢��Ƃ��w�����������܂����B���搶����́A�����n��̒������ł͂Ȃ����E�I���삩����{�̒����l���邱�ƁA�ѐ搶����́u�}�ӂ�{�ɏ����Ă��邱�Ƃ����̂܂ܐM����ȁv�Ƃ������Ƃ��w�т܂����B
�@���ށA�Ђ��Ă͍����ނ̐����n���w��i�߂Ă����ɂ͐��m�ȕ��z�E���Ԃɂ��Ă̔��\�̏ꂪ�ǂ����Ă��K�v�ƂȂ�܂��B
�@�����Ŏ��͑�w�ɓ��w����1953�N�A�����̓��u�ƂƂ��ɐÉ��������D��̌����Ɋւ��܂����B���̉�甭�s������E�u�x�͂̍����v��65���N��2017�N�ɂ�260���𐔂��A���̒ʂ��y�[�W��7000�y�[�W��˔j���܂����B
�@1957�N�t�A��w�𑲋Ƃ������͍��Z�̗��ȁi�����j���t�Ƃ��ďo�����邱�ƂɂȂ�܂����B�����āA�x�m�R�ɂ͂Ȃ����R�������Ȃ��̂��A�x�m�R�[�̑���
�͒��ޕ��z�̏�łǂ̂悤�ȈӖ������̂��A�����ē�A���v�X�ɂ�����x�j�q�J�Q���Q��̕��z�̂�����Ȃǂɂ��Ă̖���N�����܂����B
�@�������A�₪�Ď��͂���܂ł̎������̂����ɋ^������悤�ɂȂ�܂����B���Ĕ��搶���w�E���ꂽ�A�n��̒��ޕ��z�̐��E�I�Ȉʒu�Â��̖{���̈Ӗ����킩��n�߂Ă����̂�������܂���B
�@33�˂��}����1967�N�A���܂��ܐÉ���w�R�x����̂Ƃ����g�É���w�R�����r�A�E�A���f�X�w�p�������h�Ɉ�����Ƃ��ĎQ������@��Ɍb�܂�܂����B
�@�����̈ږ��D�u���W���ہi���H�j�A�A���[���`�i�ہi�A�H�j�𗘗p���đ����m��Г��łقڂP���������ēn�������̂ł��B
�@�ړI�n�V�G���E�l���@�_�E�f�E�T���^�}���^Sierra Nevada de Santa
Marta�͓�ăR�����r�A�̖k�[�̃J���u�C�ɖʂ����Ǘ��R�Q�ŁA�����̌ŗL�����������邱�ƂŒm���Ă��܂��B���͂����ł��̎R�Q�ŗL�̒������h�v�e
���������t�HMorpho rhodopteron���̏W���Ċ������A���̂悤�ȓ��X���߂����܂����i�ʐ^�R�j�B
�@����A���̎R�Q�ɐ�������n���ȃW���m���`���E�ށE�C���J�q�J�Q��Pronophilini�ɑ����邢�����̎킪�A������̂͊J���������ɁA�܂��щ�
���n�ɁA�я��̉A�n�ɁA���ɂ͗ѓ��̏��̂��������ł��ĂȂ��Ȃ��~��ė��Ȃ��ȂǁA���ꂼ�ꂪ�ŗL�̐��ԓI�n��ecological
niche���߂Ă��邱�Ƃɋ����Ռ��������܂����B���ꂱ���g��h�̂�����ł͂Ȃ����낤���B�������A�g��h�͒n���̗����ł����R�Ƃ��đ��݂���̂��B
�@���̌�A����1973�N�ɓ��{�A���f�X��c�E�����ʐM�Ў�Ấu��ꎟ���A�}�]���T���v�ɂ��Q�����A�����Ɂg�[�ԁh���߂���h�N�`���E�ނ́g��h�̂�������ώ@����@��������Ƃ��ł��܂����B
�@1968�N�A��Ă���̋A����A��Ԓ莞���Ζ��ƂȂ������́A���ĂƂȂ��Ă������{�Y�L�}�_���q�J�Q�̌����Ɏ�肩����܂����B
�@���w�Q�N���ł�����1948�N�A���͓��������É������w�Z�ɍ݊w���Ă���ꂽ���� �����璷�쌧���X�J�ō̏W���ꂽ���^�Œ��������������L�}�_���q�J�Q�̕W�{�������Ă��������܂����B���̂Ƃ�����͑��z�����z�����邽�߂ɍ�����������̂��낤�ƍl�������̂ł��B
�@���̌����͎���ɂ���Ċm���߂Ȃ���Ȃ�܂���B���͍��������g�R�n�^�h�Ɩ��邢�F���́g���n�^�h�����炵�Ă��̌��ʂ��r���܂����B�����ŗ��҂̂������ɗc���̌`�Ԃɑ傫�ȍ�������A����ɐ����̍s������j�ɂ����炩�ȈႢ�̂��邱�Ƃ��킩��܂����B
�@���̂Q�N���1970�N�Ɂu���{�Y�L�}�_���q�J�Q��Neope�ɑ������̎�ɂ��āv�Ƃ����_������{�����w��́u���Ɖ�v���ɔ��\���A���߂ē��{
�Y�̃L�}�_���q�J�Q���T�g�L�}�_���q�J�Q�ƃ��}�L�}�_���q�J�Q�̂Q�킩�琬�邱�Ƃ���܂����B���̌�A����Ԗѐ��̂������A���F�̐��̂������ȂǁA��
�̂Q��̑��݂�ۏ���f�[�^�������Ă��܂��B����͂����������Ɗ��������w�Q�N�̂Ƃ�����22�N�ڂ̔N��������Ă������ƂɂȂ�܂��B���̌����Ŕ���
�搶����g������܂��̂����t�����������������Ƃ͖Y����܂���B
�@1965�N�t�A���͐É������̕��ʎ�q���W���m�����A�쐼�����łǂ�Ȑ��������Ă���̂��ɋ����������A�����哇�E���V���ɏo�����܂����B���̌�A����
�����E���d�R�����Ȃǂւ��o�����A���{�{�y�̃q���W���m���Ƃ����̓��X�̂��̂Ƃ��������킹�Ă��̎G��̂ł����ׂ܂����B
�@���̌��ʁA�쐼�����̃q���W���m���͓��{�{�y�̂��̂Ƃ͕ʎ�ŁA�ŗL��̃����E�L���E�q���W���m���ł��邱�Ƃ��킩��܂����B
�@�L�}�_���q�J�Q�̏ꍇ�͓�����sympatric�́A�q���W���m���̏ꍇ�ُ͈���allopatric�̊W�ɓ�����܂��B
�@���̂悤�ȁg��h�ɂ��Ă̎��̊S�́A��Ăł̑̌��ɂ��Ƃ��낪�傫���Ǝv���Ă��܂��B
�@���{�Y���ނ̕��z�E���Ԃ͓��{���ӂ̋ߗ������܂ޒn��̒��ōl���A�������ʒu�t�������Ă����K�v������A�����Ă˂Ɂg��h�Ƃ͉������ӎ����Ă��̕�
�z�̊g��o�H�Ȃǂ𐄒肷��B���̂悤�ȍl��������A����1970�N�ȍ~�A��p�E�����嗤�E�؍��E�����S���E�����A�W�A�i�L���M�X�j�E�ɓ����V�A�Ȃǂ֏o
�����A�����ɂ����̒n��ɂ�������{�Y���ނƂ̋��ʎ�₻�̋߉���̕��z�E�����n�E���ԂȂǂ����ۂɒ������邱�ƂɂȂ�܂����B
�@�����̊C�O�����ɂ����āA���̓`���E�Z���q���q���E�����A�E���O���z�V�~�X�W�A�^�C�����L�}�_���q�J�Q�A�E���i�~�W���m���ނR��ȂǂɊւ��āA�g��h�ɂ��Ă̒��s�Ȃ��܂����B
�@���N���ォ�狻�������������Ă����A���v�X��x�m�R�̒��ޕ��z���ǂ̂悤�ɂ��Č`�����ꂽ���Ƃ������Ƃ��������킩���ė����悤�ȋC�����܂��B
�@��l�I�̕X�͊��ɂ����鐼����̗≷�уX�e�b�v�A�k����̊��уc���h���̓��{�ւ̐N���A��X���i�q�v�V�T�[�}�����j�̉��g���ɂ��A���ω��ȂǁA���{�̒��ނ͊��ω��̑傫�Ȏ������o�č����̏�Ԃ��`�Â����Ă�����̂ƍl�����܂��B
�@���ނ̕��z�͂��́g��h�̐����̗��j�Ɛ[���ւ���Ă���ɂ���������܂���B�킽���͂��̖��̉𖾂ɗ]������������ł��B
 �@�@�@ �@�@�@
�i�ʐ^�Q�j�@�@�@�@�@�@�@�@�@�@�@�@�i�ʐ^�R�j
|
���� �^�|
�@
�������R�j���l�b�g���[�N����
(�ʐ^�P�j
|
2018.1.19
No.24�@�u�n������W���n���������v
���́A���͎��ɂ��邱�Ƃ��w�ǂȂ��Ȃ����u����
�w�Z�v���w�O���璎�D���̍������N�̂Ȃ�̉ʂĂł��̂ŁA��������́u�����v�ł��B���Z���̍��͓��{�̑�w�ł̍����w�̋���Ȃǂɂ��Ėw�ǖ��m�ł�����
�ŁA�����P�ɎG���A���ɏ��a21�N���́u�V�����v�Ȃǂ̒m������A���{�ł͍����͖k�C����w�Ƌ�B��w�A�߂��ł͓����_�H��w�������̌�����ɂ����
���邭�炢�̒m���ł����B�R�����b�{�s�ň�������͓��������ł����w���������Ă����k�C����w�́u�g�߂��v�ŁA�������b�{�ꍂ�̐������̌E�c�搶���k�傲
�o�g�ł����̂ŁA�k��̗��Ȃ����悤���ƍ��Z3�N�̑O���܂ł͌��߂Ă��܂����B�Ƃ��낪�A������R�̂��������ŋ�B��w�_�w�������邱�ƂɂȂ�A
�K�^�ɂ����i���āA�ȗ��S�O���N���̑�w�Œ���̐����������܂����B2000�N�ɑފ�������A80�ɂȂ鍡�������̒n�Œ����������Đ��������Ă���
���B
�]���O�搶��������O�搶�̂��Ƃŋ��_�w�������w�����̑�w�@�C�m�ے����łĂ���A���a36�N�ɕʃL�����p�X�̋��{���̐����w�����̋����ɂȂ�A����
���A�{�{����̗��搶���ݐE����Ă��������w�����Ő����w�̈�ʋ�����Ƃ߂܂����B�������g�̌����̓~�m�K�ȁi�c�����~�m���V�j�̌n�����ފw����A����
�ɑo���ށA���ɃI�h���o�G�Ȃ̌n�����ފw�ɏd�_���ڂ��Ȃ���A�����搶�̂���`���Œ��ނ̌����������Ă��܂����B���{����199x�N�ɉ�̂���āA�V����
��r���������ȂƂ��������w�ۂ̑�w�@�̒n�����R���u���̋����ɂȂ�A���^����A��c�������Ƌ��ɏ��߂āu���O�v�̑�w�@���������邱�ƂɂȂ�܂�
���B���̑O�����͔_�w�������w�����̊w����@���Ƃ͌����ʂŐ[���֘A��ۂ��Ă͂��܂����B�u���̏��߂Ẳ@���ɂ́A�O�O��w���_���R�ώ@���̒������V�N�A
���i�Δ����ق̖��i��G�N�A�k�卩���̌n�̋g�V�a���N�A���A�Z�X��Ђ����c���Ă����䐽�N�炪���܂����B
�ނ�͑o���ځA�`���^�e���V�ށi���{�ځj�A�����ڂ̌n�����ނ��C�m�_���̃e�[�}�ɂ��Ă��܂����B�{�{�搶����̍����`�Ԋw�d���̎��́A�ނ�@���ɂ͂܂���
�ۂ̍����ނ̏ڍׂȌ`�Ԋw�I���������邱�Ƃ����߂܂����B����͒P�Ȃ�����̕��ފw�ł́u���ތ`���v�ł͂Ȃ��A���h�C�c�̑o���ޕ��ފw�҂̃w���j�b�N
�iWilli Hennig�j������n���̌n�w�iphylogenetic
systematics�j��ڎw����`�Ԋw����������悤�ɁA�Ƃ����|�ł����B�����ƍ��C�̂���w��Ɩڂ��g�����C�Ə����̎d���ɂȂ�܂��B�D�G�Ȕނ��
�������҂����悤�ɗD�ꂽ�����𑱂��Ă��܂����B�V������w�@���ł����͓��{�ł����V�搶�Ȃǂ��I�T���V�Ȃǂ�Ώۂɂ��āA�悤�₭�����镪�q�n���̌�
�����n�܂肾�������ł����B�������A���ނɂ����������������́A�M�t�`���E�ނ̐i���ɂ��S���[���A�`�Ԋw�I�Ȍ����𑱂��Ă��āA�Ĕ������ꂽ�����l���
�̃E���i���V�{���A�Q�nBhutanitis
mansfieldi�̌n���I�ʒu�Ȃǂɂ������������Ă��܂����B�������A�n������W�̐���ɂ����āA���̔\�͂������Ă��Ă͌`�Ԋw�I��@�ł͂킩���
�����������邱�Ƃɂ́A��X�C�t���Ă��܂����B���̕����́A�c�������܂߂��`�Ԋw�ɉ����Ă̍s���w���܂ސ��Ԋw�A�����n���w�A���X�̒m���ƂƂ��ɁA���q�n
����͂���̉�����i�ɂȂ邾�낤�ƁA���V�搶�̃I�T���V�ނ̌����Ȃǂ�ʂ��Ċ����Ă��܂����B����ŁA�C�m�ے��ŃA�V�i�K�o�G�Ȃ̌��������Ă������i
�N�ɁA�M�t�`���E���Ƃ��̎��ӂ̕��q�n����͂�����������ǂ����ƒ�Ă��܂����B�ނ��n���I�핪���̌����ȊC�l���̃A�V�i�K�o�G�̌��������낻��ڎw����
�������Ƃ��A�ނɊ��߂����R�̈�ł����B���Ƃ͑S��������āA�������q�n����͂̎������ł���ꏊ�͂��܂葽���͂Ȃ��āA���V�搶��h��������
��Ă���JT�����������ق̌��������K���ł��낤�ƍl���āA�ނɊ��߂܂����B�K���A�����������ق����i�N������Ă��������āA�����ŏ��߂Ď������̍u
�������q�n����͂ƊW�����Ă�ɂȂ�܂����B
�@���́A�W����@���ɂ́A�����ΏۂƂ��Ă܂������̎��ԁA���Ɍ`�ԍ\���͍������������̒��Ő����Ă��������ŁA���ڊO�E�ƑΛ����鑤�ʂ������Ă����
�ŁA�`�Ԋw�I�����A���ɊO���`�Ԋw����ɓO��I�ɍs�����Ƃ��������āA�����ʂ��Čn����͐��_�ɂ��Ă��n���_�ɂ��뉽�炩�̌��ʂ����������ŁA���q�n
����͂��s���悤�ɁA���Ȃ苭���������Ă��܂����B���i�N�������������قŌ����̏����̌P��������������ɁA�������̑�w�@��DNA����z��͂ł���
�u�����ł��܂����B���̍u���̏��r�T�q�����̎w���ŁA���i�N�ȊO�̉@�������q�n����͂ւ̃A�v���[�`���n�߂܂����B�����āA���̌㎄�����̍u���̊W�҂�
�ł��傫�ȉe����^���ĉ��������̂��A��b�����w�������̖ї��G�Y�搶����ђ��J�����א搶�ł���܂����B
�ї��搶�͂����̌����̂ق��ɍ����A���ɒ��ނɐ[���S�������Ă����āA���ނ̌n����͂��n�߂��Ă��܂����B�����āA�J��R�I�q������Ƃ��Đ�
���̌����̂���`��������Ă��܂����B������DNA����z���͂̎����̏K���ƒ��ނ̕��q�n����͂̌����̂��߂ɁA�������̍u���̉@�������X�Ƃ����b�ɂ�
�邱�ƂɂȂ�܂����B�������̑�w�@�̃X�^�b�t�ł��������ތ����҂̖�c������A�@���̏��c�،���N�i������r�����������������j�A��㏟��N�i������
��w�������������ُ����j�A���{���،N�i���^�ߍ������A���~�n�r���ِ����j�A����Ɏ����g������̌������ɑ؍݂����Ă��������āADNA�̕��q��͂�
�C���n��������Ă��������܂����B�����ł̌o���͌�ɔނ�̍����n���w�̌����ɑ傫�ȉe����^���邱�ƂɂȂ�܂����B�܂��A���r��������DNA��͂̎�
�@��������g�V�a���N�͖k�C����w�ɐE�āA�����Ŗ{�i�I�ɍ����ނ̎�v�Q�A���ɕs���S�ϑԗނ̌n����͂������Ȃ��ƂƂ��ɁA����獩���ނ����̊
�̔��ׂȌ`�Ԋw�I������{�i�I�Ɏn�߂�悤�ɂȂ�܂����B
����܂Ŏ����߂��鍩���̌����A���Ɍn���w�I�����ł́B�`�Ԍ`�����d�����Ȃ���V����������i�Ƃ��Ă̕��q�n����͂֒��ڂ��Ă������Ƃ�����������������
���Ǝv���܂��B�ŏ��ɏ����܂����悤�ɍ�������̒��D���ł����A���̗l�X�Ȗʔ����̒��ł����낢��ȍ����̈Ⴂ�A��r����A���̋N���A������T�������Ƃ�
���_�ɋ������ڂ��čs���܂����B���A���w���̍��͐��c�ɐݒu���ꂽ�U�铔�ɏW�܂������������āA�^�K����Q���S���E�A���܂��܂ȉ�Ȃǂ��낢��ȍ������W
�߂Ċ��ł��܂����B���Z�ł͂����ς璱�ނɋ������ڂ��Ă��܂������A��w�ł͔_�w���Ƃ������ƂŃ~�m�K�Ȃ̌������n�߂����ƂŁA���̒��B�̎��̒�������
�l���̋N����m�肽���Ƃ����C�����������Ȃ�܂����B����ɁA�����A�����搶�ƎR�{�p�䂳�~�h���V�W�~�ނ̌n���w�̌��������Ă��܂����B���̂悤�ɂ�
�낢��ȉe���̂��ƂŁA���������̋N����T�肽���A����Όn���w�I�Ȍ������u������悤�ɂȂ�܂����B
���s��w�Ńl�Y�~�ނ̌����Ȃǂ����ꂽ���c�䖫�搶��������g�S������u���e�i���_�v�Ƃ����{���o����Ă��܂����B����͊w�����̎��ɂƂ��Čn���w�X��
���Ă������߂̑傫�ȓ��W�ɂȂ�܂����B�`�ԂƐ��������ڂɊ֘A���Đi�����Ă���~�m�K�̌������n�߂Ă����̂ŁA�n���w�͐����̐�����ʂ��Č���������
�̂ł���A�Ƃ������c�搶�̖���͋����S�ɍ��܂�܂����B�����āA�n���w�̏d�v����[�I�ɂ��߂����}���A�w���j�b�N�iWilli
Hennig�j�������� Phylogenetic
Systematics��1966�N�ɏo����ƂɂȂ�܂����B���̖{�͌n���̌n�wsystematics�̏d�v���A�n���}����T���̎�@�A���̏d�v����
�����ɁA���������Ȃ����_���Ŏ�����Ă��܂����B�{��̏��_�ł͓N�w�I�Ȍ������W�J����Ă��āA�ُؖ@�I�B���_�ɊS�������Ă������ɂ͂܂��V�N�Ȗ�
������܂����B���ɂ��Ă݂�ƁA�{���̓ǎ҂̂Ȃ��Ŋ��l�̕��X�����̖{�̖��O�͒m���Ă��Ă��A�ǔj�����ł��傤���Ɩ₢���������C�����ɂȂ�܂��B�w��
�j�b�N�́A�n������̌n�͐i�������w�ɂƂ��čł��d�v�Ȉ�ʎQ�Ƒ̌n�igeneral reference
system�j�ł���A����䂦�ɐ����w�����̒��ł��ɂ߂ďd�v�Ȍ�������ł��邱�Ƃ��������Ă��܂��B���̂悤�ɁA���̌n���w�́A�~�m�K�⏔�X�̍�����
�����瓾�����̒m���Ɠ��c���̌n���w�A������Hennig�̌n���}�T���Ƃ�����{�̓��Ői��ł����Ƃ������Ƃ��ł���ł��傤�B�����܂ŁA���Ǝ������
�����n���w�W�̎����̈ꕔ���q�ׂ����Ă��������܂����B
���āA�悤�₭�{����Ȍ��ɏq�ׂ����Ǝv���܂��B�����̓�̑��ʁA�`�Â���Ɠ����Ƃ����ϓ_����́A����DNA�́u����z���ԁv�͑O�҂ł���A������
�̌`�Ԃ��̂��̂ł��B���̓_���\���Ɏ��o���ꂸ�ɁA�����̌`�Ԃ�DNA�z���Δ䂳���āA�����������҂��ʌ̐����̑��ʂł��邩�̂悤�Ȍ�����l������
�����҂������ɑ������Ƃ��B�����̌`�ԂƂ͕��q���x�����獩���́u�O���`�ԁv�܂ŁA���B���Ƃ炦�悤�Ƃ���ΘA���I�ȃ��x���̂����ꂩ�ł���܂��B���
�̊ϓ_��DNA����z��͌`�Ԍ`���̈�ł���Ƃ������Ƃł��B�������DNA�������I�ɋ@�\����ʂ͕ʂł��B
���ɁA�w���j�b�N�͓����̂Ȃ��Ōn������W�̒m���͈�ʎQ�Ƒ̌n�ł���A�����̑��l�ȑ��ʂɂ��Đ����̊ԂŔ�r�����Ȃǂ���ۂɎQ�l�ɂȂ�̌n�Ƃ���
�@�\���邱�Ƃɑ��݈Ӌ`�����邱�Ƃ����������Ă��܂��B���Ȃ킿�����̊O���`�ԁiintegumental
morphology�j�̔�r�����ɂ���Ď�Ȃǂ̊Ԃ̗މ��W�𐄘_����ɂ��Ă��A�܂�DNA����z��Ɋ�Â��ē��l�ɗމ��W��}�ɕ\���Ă��A��
���������ʎQ�Ƒ̌n�����グ�錤���ł���A�Ƃ������Ƃł��B���̊ϓ_�́A�n������W�͔�r�����w�A���邢�͐i���w�A�n�������w�̂��߂̈�ʎQ�Ƒ�
�n�ł���Ƃ����_�ł��B
��O�ɁA�n���w�̌����������ŁA���t�̒�`���d�v�ł���Ƃ������Ƃ��q�ׂ����Ǝv���܂��B��X�͌��t��ʂ��Č����������������g�̒��ł��A�����ҊԂł�
�F���������Ă��܂��B�n���A�n���}�A�n������W�A�n�������A�n����́A�ȂǗl�X�Ȍ��t�������҂̒��ʼn��C�Ȃ��g���Ă��܂��B���X�̌��t�̒�`�ɑS��
�G������͂���܂���B���ł��A�ł����R�Ǝg���Č�����₷�����t���u�n���v�ł��낤���Ǝv���܂��B�l�ɂ���Ăǂ��ɂł��̂��Ă��錾�t��
���B�n������W�́A�O�ȏ�̐����킪�ߋ��Ɍ����������ꂽ�����ł���A�ߋ��̎����ł����āA���B�͂���𐄘_���邵������܂���B�����Ɍ�����������
�͂����܂ł����肳�ꂽ�n������W�ɉ߂��܂���B�ߋ��̌����Ɛ��_�ꎋ���Ă͏�ɂ����Ȃ��̂ł��B�������n������W�́u�n���v�ł͂���܂���B
��O�̊ϓ_�͌n�������̌�����ł͌��t�̒�`�A�T�O�m�ɂ��Č������ׂ��ł���Ƃ������Ƃł��B
�Ō�ɁA�n������W�ƌn�������ł��B�w�b�P���iHaeckel�j�́u�̔����͌n������������v�ƊȌ��ɂ܂Ƃ߂��Ă��锽�����ł��B���������̂�
�̂������ʼn]�X����̂ł͂���܂���B�����ł͌̔����Ƃ������t�ƌn�������Ƃ������t���Δ䂳��Ă��܂��B�̔����ƌ����A����ł���������q��َ�
�̔������v���o���܂��B�������A�����Ȃǂł͌���q�����Ō����ȕω��������܂��B�����āA���S�ϑԗނ́u���A�c���A匁A�����v�̌o�߂����ǂ�ƒP��
�ɐ}���I�Ɍ����Ă��܂��܂��B�������A���ꂼ��̎�ł́A�����c����匂�������ꂼ��̎p�������܂������Đ��������Ă��܂��B���̕ω��ߒ����̂��̂�
�����ȈӖ��ł͌̔������Ǝv���܂��B�����āA�����n���������̔����̔����ł���A�Ƃ���A����͍����鐶���̂�����A�`����A�́A�ߋ��ɑ��݂���
�c�悽���̂�����A�`����̘A���̒��ɂ���A�Ƃ������Ƃ͎����̂��Ƃł��B�n�������Ƃ́A�n������W�i�����ɂ͐��肳�ꂽ�j���̂��̂ł͂Ȃ��āA���
���܂����A�ߋ��̑c�悽���̐������܂̘A���ɑ��Ȃ�Ȃ��Ǝv���܂��B������������Ă����̂��A�n�������w�ł͂Ȃ��ł��傤���B���A�R��ȏ�̐����̍���
�ߋ��̑c�悽���̎p�Ɛ�����T������A����Όn�������������E���_���āA�܂��A�R��̐������H���������n��A�����̌n���}�̕���W�̕�����}������
�A���ꂪ�n���}�}�ł���킯�ł��B����͑O�q�̂悤�Ɍn���������̂��̂ł͂Ȃ��A���̈�ʂɂ����܂���B�������ADNA����z��ł����炳�ꂽ����}�A
����Ε��q�n������}���A�u�n�������v���̂��̂ł���悤�Ȍ��z������Ă��錤���҂�����悤�ȋC�����ĘV�w�͐S�z�����Ă��܂��B���q�n������}��
Hennig�̌����Ƃ���̈�ʎQ�Ƒ̌n���\�z���邽�߂̈�̎����ɂ����߂��Ȃ��̂��Ǝv���܂��B�Ō�Ɍn�������͌n������}�ł͂Ȃ��Ɛ\���グ�āA�I
���܂��B
|
�O�}�@ �L��
�@
��B��w���_����
�O�}�������R�j������
|
2017.4.28
No.23�@�u���E�̉ʂĂ܂ōs���č����̏W�@in �C���h�m�v
�C�݂ō����̏W�B���̑g�ݍ��킹�ɂ��܂�����
���Ȃ����Ǝv���܂��B���̌����Ώۂ͒��ԑтɐ������Ă���A�V�i�K�o�G�̒��Ԃł��B���̃C�\�A�V�i�K�o�G�͒��̖�������������C�݂Ő������Ă��܂��B���H
���̃n�G�ŁA�c���Ɛ����͐ߑ�������ߐH���܂��B���A�c���A匂̎����ł́A���������Ă���Ԃ͊C���ɒ���ł���A��������������������Ă��钿���ł��B��
�̑��ɂ��A���l�̃J�j���ɂ���n�G�����܂��B������͐��Ԃɂ��Ă͕������Ă��܂���B�����̍����͊C�݂̒���ɂ��������ł����A���z�悪����ƂȂ邽
�߁A�n�������n���̍ޗ��Ƃ��čD�s���Ȃ̂ł��B�C�m�_���ł͓��{�A���m�_���ł͓��A�W�A�n��Œ������Ă��܂����B���݂ł͒����n��𐢊E�ɍL���Ă���
���܂��B
�@�n���͂����鏊�ɒ������邱�Ƃ���u���̘f���v�Ƃ������܂��B�Ƃ͂����Ă��A�C�݂ɂ��ލ����̎�ނ͑����͂���܂���B���̂��߁A�C�݂ł̍̏W����
�����Ԃɗ��߂��A�����O�̔����قɂ��ړ��Ă̕W�{�͂���܂���B�����ŁA���͂Ńn�G���W�߂邱�ƂɂȂ�A���܂łQ�X�̒n��⍑�ō̏W���Ă��܂����i�}
�P�j�B�A�V�i�K�o�G�̒��Ԃ͖��L�ڎ킪�����A����̏W�����n�G�ɂ܂����O�������Ƃ���n�߁A�����𑱂��Ă��܂��B���m�ے��P�N���̂Ƃ��ɑ��V�ȎO�搶
�̂����ӂőh�q�d����̂��Ƃ�DNA�̎������@���w���Ē����܂����B
�@�����Q�N�قǂ̓C���h�m�ɏœ_�����ĂĂ��܂��B��N�Q�`�R���ɂ́A�}�_�K�X�J���A�}���b�g�A�����j�I���A���[���V���X�ŁA���N�̂R���ɂ̓A���r�A������
�I�}�[���ƃJ�^�[���A�Z�C�V�F���ƃ����f�B�u�Œ������܂����B��������u�y���ō����̏W�v�Ɏv���܂����A�����Ŋy�������ɉj���l������A��ނ������l��
�������ڂɁA���A�̂܂������Ȃ��A���˓����ɏƂ炳���ܔM�n���̒��ň�����A�����Ԃ��ӂ葱���Ă��܂��B�w������ɓ����������̃t�B�[���h�ɂ͊���Ă�
��F�l�ƈꏏ�ɊC�݂ō̏W�������Ƃ�����܂����A���̗F�l�͈���łЂǂ����Ă��ɂȂ��ςȂ��ƂɂȂ�܂����B���Ă��Ɏア�l�͗v���ӂł��B���͂Ƃ���
�ƁA���߂Ă̏ꏊ�ł����n�̐l���瓹����邱�Ƃ��x�X���邭�炢�A���n�̐l��荕���Ȃ�̂ő��v�Ȃ̂ł��B
�@���̒������@�͒����ꏊ�Ɠ��l�ɑ��̍��������Ƃ͈Ⴂ�A�P�ӏ��ɍ��𐘂��邱�ƂȂ��A�]�X�ƈړ����Ă����܂��B����C�ݐ��łǂꂾ�������̒����n�_����
���邩�ɂ������Ă��܂��B���̂��߁A�����n�̑I������O�ɂ������肵�Ă����Ȃ��ƁA���n�Ŏ��Ԑ�ɂȂ�A���̒����n�Ɉړ����Ȃ��Ă͂Ȃ�܂���B����
�ŁA���O�̒����n�I�肪�d�v�ƂȂ�܂��B�ǂ̂悤�ɂ��Ă��邩�Ƃ����ƁA�O�[�O���}�b�v�̉q���ʐ^���g���A�C�\�A�V�i�K�o�G���������Ă������Ȋ��̗L��
���m�F���Ă��܂��B����ɂ��A�قڊԈႢ�Ȃ��ړI�̍������̏W���邱�Ƃ��ł��܂��B�֗��Ȏ���ɂȂ������̂ł��B�܂��A�t���J�嗤�ł͓�A�t���J�ƃ�
���b�R�ł����������ł��Ă��܂���B�����n���^�[�̃��[���h�c�A�[�́A�������������܂��B
�}�P�����n�_
|
���i�@��G
�@
���ꌧ�����i�Δ�����
���w�|��
|
2017.4.28
No.22�@�u�N����T���ăn�`�����v
�R�����̃o�g����n���Ă�������������搶�̌�
�����́A���ɑ��l�ȕ��ތQ��ΏۂɌ�����W�J����Ă��Ċ��Q���܂������A���̏o�g�̌������ł��w���͂����Ԃ�Ɨl�X�Ȍ����Ώۂ������Ă��܂����B����ȉ�
�̐[���̂������ł��傤���A������]�ʂ�q���o�`�Ƃ����o�`��ΏۂƂ��ĕ��ނ̌������X�^�[�g�����邱�Ƃ��ł��܂����B�Ȃ���₱�������ȃq���o�`��
�̂��Ǝ����ł��v��Ȃ��ł��Ȃ��̂ł����A��v���o���̂́A���������̎��K�ŏ��߂ăX�E�B�[�s���O�ō̏W���������́A�F�`�̗l�X�ȃq���o�`����R�Ԃ�
���������i�ł��B�������ׂĂ݂����̗̂e�Ղɂ͐��̂��킩��Ȃ��A����Ȃɂ悭������Ȃ������g�̂܂��ɂ�������ƌ����̂͐V�N�ȋ����ł����B��
���A�����������獩���A���ł��n�`���D�����������߁A���R�ώ@�҂̎�L�Ȃǂ̊�c�v��Y����̒���͏�����������悭�ǂ�ł��܂������A�N���Ɋ���N
���q���o�`���͂��߂Ƃ��āA�������̃q���o�`�����̒��ň����Ă��āA�q���o�`�̍s���͖ʔ��������Ƃ�����ۂ��������̂����R�̈�Ƃ����܂��B��w�@
�ł͍ސ��E���̍b���Ɋ���q���o�`�̕��ނ�A�g�K���q���o�`�̐����j�̌����Ɏ��g�݂܂����B
�@���̌�A�^�ǂ��w�|���Ƃ��Ĕ����قɐE�܂������A�w�|���̎d���͎����̎��W�ۊǁA���������A�W���A���y�E����Ƒz���ȏ�ɑ���ɂ킽��܂����B����
�Ƃ̃o�����X���Ƃ�̂��Ȃ��Ȃ��e�Ղł͂���܂��A���Ԃ��ʂ��Ď��g��ł���̂��N���q���o�`�̌����ł��B��͂�s�����ʔ������Ƃ����̂��傫�ȓ�
�@�ƂȂ�܂����B�N���q���o�`�͊O���ߐH�����E���Ƃ���������ƕς�����l���������Ă��܂��B���X�͊��ƂȂ�N����j�Ŏh���Ė������Ă��痑��
�̕\�ɎY�ݕt���܂��B�N���ւ̃A�v���[�`���Ԃ̎�����щ���Ă����т�������́A�ԂɂƂ܂��ăN�����߂Â��Ă���̂��Ђ�����҂��́A����I�Ԃɔ�
�э���A�q���O���ނ̒ނ莅���r�ł͂����āA�l�����Ǝv���Ĕ�яo���Ă����N���ɋt�P������̂ȂǂƂĂ����l�ł��B��������o�߂��N���͒ʏ�̐�����
�߂�A�z�������n�`�̗c���͂��̑̂̏�ŏ������̉t���z�����Ȃ��琬�����Ă����܂��B�����A�����̉^��������E������w�����A�������ƖԂ�A�l����
�߂炦�鈣��ȃN�������I
�@������N���q���o�`�Ŗʔ����̂̓n�`�̗c���ɂ��N���̍s������ł��B�N���̖ԂƂ����̂͊l���̕ߊl�ɔ��ɓ������Ă��܂����A�����Ԃɂ킽���v��
�ɂ͌����Ă��܂��B�N�������C�Ȃ����͖Ԃ͏�Ƀ����e�i���X����Ă��܂����A�N���͍ŏI�I�ɋz���s������A����ł��܂��̂ŁA�Ԃ̏�Ŗa�����}���́A��
�͂��Ȃ�댯�ȏɒu����邱�ƂɂȂ�܂��B�ł͂ǂ�����ăN���q���o�`�͂���Ȋ댯�ȏ������̂ł��傤�B�s�v�c�Ȃ��Ƃɏ\���ɐ��������c����w
�������N���́A�Ȃ����s�����ω����A��菬�����A�c�����d�����A�S����������������������v�ȖԂ�悤�ɂȂ�܂��i�}2�j�B�c����������ʂ��ĉ���
���̕����𒍓����Ă���ƍl�����Ă��܂����A�����ɑ�����悤�Ɂi���ۂɑ����Ă��邱�Ƃ��c���̎�菜���ɂ���Ċm�F����Ă���̂ł����j�A������
����ɌJ��Ԃ�����⋭���Ă����p���݂Ă���ƁA���Ƃ������Ȃ��C�����ɂȂ�ƂƂ��ɁA�����̑��ݍ�p�̖��ɐ[�����S�������܂��B�������Ē��点��
���S�ȕό`�Ԃ̏�ŁA�N���q���o�`�̓}����a���A匉��E�H�����Đ����ƂȂ�܂��B
�@�N���q���o�`�����̂Ƃ肩����Ƃ��ċL�ڕ��ނ�i�߂A�ǂ�ȃN���𗘗p���Ă���̂��A�c�����A�����̍s���A���̑���ɂ��āA��O�ώ@�⎺������
�Œ��ׂĂ����܂����B���{�����50��ȏ�̃N���q���o�`���������Ă��܂����A���p�����N���͗l�X�ȃO���[�v���܂݁A�قƂ�ǂ̏ꍇ�P��Ȃ����A����
�߉��Ȑ���̃N���𗘗p���Ă��鎖���������Ă��܂����B�N���Ƃ̌��т��̋������l�����������R�Ƃ������Ƃ���ł��傤���B�Y���s���A��Ԃ���ю��
�̋����ɂ��Ă��ώ@��𑝂₵�A��呀��ɂ͖Ԃ̋��������łȂ��A�ߐH�҂ɑ���h��̋@�\�����邱�Ƃ����炩�ɂ��邱�Ƃ��ł��܂����B
�@���ꂼ��̃N���q���o�`���ǂ�ȃN���𗘗p���A�ǂ�ȁu�Z�v�������ăN�����U�����Ă���̂��A���̉��̂ƂĂ��y�������̂ł����A��W�܂��Ă�
��ƁA���x�͂��ꂼ��̃N���ƃN���q���o�`�̊ւ�荇�����ǂ̂悤�Ɋl������Ă����̂��A���̗��j��`���Ă݂����Ȃ�܂��B�T���v���͂��łɑ����Ă��܂�
�����A���傤�Ǘǂ��^�C�~���O�ŊقɃV�[�N�G���T�[�ƃT�[�}���T�C�N���[����������܂����̂ŁA������͂ɂƂ肩����܂����B���̌��ʁA��܂��Ȋ�嗘�p
�̓��A�܂�N���q���o�`�̓N���̗��X���̗���H�ׂ�O���[�v�Ǝo���Q�W�ɂ���A�t�N���O���̂悤�ȖԂ炸�ɉB��Ƃ����O���[�v�𗘗p�������
���܂����A����ɃA�V�i�K�O����T���O���̂悤�Ȑ����~�Ԃ�M�Ԃ���̂𗘗p����O���[�v�ƁA�I�j�O���̂悤�Ȑ����~�Ԃ�N���̊҂ɕ���
�������Ɓi��҂̈ꕔ�͂���ɕs�K���ȗ��̖Ԃ�q���O���ނɃV�t�g�j�����炩�ɂȂ�܂����B
�@�����̒����ł͂��ꂼ��Ώۂɂ���ăT���v�����O�̕��@�͈قȂ�܂��B�N���q���o�`�̍̏W�͖Ԃ��g�����X�E�B�[�s���O��}���[�[�g���b�v����Ȃ��̂ł�
���A�����邽�߂ɂ͗c���̂����N����������̂���Ԃł��B�Ԃ������ɃN���������āA����c�������Ă��Ȃ�����������`�F�b�N����̂���
�{�ł��B�܂��r�[�e�B���O���������Ȃ��̏W�@�ŁA�l�b�g�����Ɏt��}��@���A�����Ă����N�����P�����`�F�b�N����Ƃ�����T�������X�ƌJ��Ԃ���
���B���ł����啔���̎�̗��p����N�����������Ă��܂����A�������n�߂������́u���̃N���ɂ��c�������Ă���I���̃N���ɂ��I�v�Ƌ����̘A���ł����i�}
2�j�B�c���t���̃N���͂�������A��A�n�`���H�������邽�߂Ɏ��炵�܂��B�q���O���ނ͋����e��ł����炪���܂��������Ƃ������̂ł����A�X�y�[�X��
�W���I�j�O���Ȃǂ͂Ȃ��Ȃ����܂������܂���B�����߂Ċm�F���ꂽ�N�����r���Ŏ���ł��܂��͖̂{���ɂ�������ł��B����ł��A�ȑO�͉t�Z�W�{�ɂ�
�ďI��肾�����Ƃ��낪�A���͗c������DNA�𒊏o���ăo�[�R�[�f�B���O�Ŏ�̐�������邱�Ƃ��\�ł��B�قȂ�X�e�[�W�Ԃł���r���邱�Ƃ��ł���Ƃ�
���̂�DNA�̑傫�ȋ��݂ł��ˁB��������������ēǂz�S���m���Ă��Ȃ����̂������Ƃ������Ƃ�����A���̏ꍇ�͂قږ��L�ڎ�ł���Ȃ���A��
���Ƃɂ͎��c�������c�炸�A�����̎p�`���S���킩��Ȃ��Ƃ����A����͂���Ŕ߂����Ɋׂ��Ă��܂��܂��B
�@�����̎�Ŋ�傪�������Ă��Ă���Ƃ͌����A�܂��܂����̕s���ȃN���q���o�`�͎c���Ă��܂��B����܂ł̔����̃y�[�X���l����ƁA���m�̃N���q���o�`
���܂��m���ɂ���ł��傤�B�P��̎�Ǝv���Ă������̂ɁA�������̈�`�I�Ȃ܂Ƃ܂�̂���A���ꂼ��Ⴄ���𗘗p���Č`�Ԃ������ɈقȂ��Ă���W�c
���F�߂�ꂻ���A�Ƃ����Ⴊ�������̃O���[�v�ŕ������Ă��Ă��܂��B���܂��t���̃N���T���͂܂��܂��I��肻���ɂ���܂���B

�}1�@Reclinervellus tuberculatus�̗c���̑���ɂ��ό`�����S�~�O���̖ԁi���Ƃ͐����~�ԁA�����̓}���j

�}2�@�z�V�~�h���q���O���Ɋ���Zatypota chryssophaga�̗c��
|
���{ �����Y
�@
���s�����R�j������
�w�|��
|
2017.2.20
No.21�@�uDNA�̃p�Y���v�Ɓu��炵�ג��ҁv
�uDNA��͂̓p�Y���̂悤�Ȃ��́v�B�w���ɐ�
������ۂɎ����悭�g�����t�ł��B�z��߂���A�n�������������A�v���C�}�[��v������A��@������߂܂Ŏ��ɂƂ��Ă͓���y�����p�Y���̘A����
�����B�X�����Ɛт͂Ȃ���������܂��A�����ł͂���܂ł̉ߒ��́u��炵�ג��ҁv�̂悤�ȗǂ��@��̘A���������Ǝv���܂��B
�@�w�ʌ����ł́A�A�T�M�}�_���Ɗo�G�̎�ԊW�𒆐S�ɖ�O�����Ǝ���������s���Ă����̂ŁA���̔��m�_���ɂ�DNA��͂͑S���o�Ă��܂���B�����A
�����ň������������̈�`�I�Ȕw�i�͂����C�ɂȂ��Ă��܂����B2003�N���낾�����Ǝv���܂����A�����L�����p�X�̔��؍F�i�搶�ɂ����b�ɂȂ�ADNA
��͂̋Z�p�������܂����B�}�_���`���E��o�G�́A�O���ł͕�����Ȃ��Ⴂ�����������Ƃɑ�ςȊ������o���܂����B�قǂȂ����̌������ɂ��Œ��
PCR���ł����ނ�����A���������ŃV�[�N�G���T�[�����p�ł���悤�ɂȂ�܂����B���̃^�C�~���O�ŁA�V���r�A�V�W�~�̑��̌Q����������A���g�̌�
�����ł̉�͂̑����ƂȂ����̂ł��B���߂Ĕz���ǂ̂́A�����ł͂Ȃ��A�����ۂ�Wolbachia�ł����B���̌����́A�O��R�����̍�{���q��
��ɂ���Ĕ��W���Â��A���܂��܂Ȕ����ݏo���܂����B2008�N�ɖ{�B�ő唭�������N���}�_���\�e�c�V�W�~�̉�͂ł́A�������������̂��܂߁A
���܂��܂ȋ����̑����̕W�{���������̂ŁAPCR�̏���������ǂ����ɂ��Ȃ�܂����B
�@���ɂƂ��Ă̎��̃X�e�b�v�A�b�v�́A2010�N�̃t�B�������h�ւ̗��w�ł����B�O���́A���^�̌Q�����ŗL���ȃO�������B���q���E�������h�L�̐����n
�I�[�����h�����ŁA�V�G�̃R�K�l�R�o�`�̖�O�����Ǝ���������s���A���̌�w���V���L��w�ŏ��߂Ẵ}�C�N���T�e���C�g��͂��s���܂����B�����A�����O
���[�v�ł̓O�������B���q���E�������h�L�̑S�Q�m����͂Ɏ��g��ł���A�����̌����ҁA�w���A�Z�p�҂�����Ɍg����Ă��܂����B�V�[�N�G���T�[������
�ςȂ��ŁA���̃T���v��������]�n���Ȃ���Ԃł������A���x�����荞�܂��Ă�����ĉ��Ƃ��f�[�^�邱�Ƃ��ł��܂����B
�@���̌o���͂̂��ɁA�W�����ނ̃i�K���z�g�P�h�W���E�̌����Ŗ𗧂��ƂɂȂ�܂����B�M�S�Ɍ������s�������˕j�y�N�̓w�͂ŁA���Ƃ��Ƌ��ނ��D�����������ɂƂ��Ė����������̘_������N�o�ł���܂����B
�@���̌��������l�����܂����B�������ł́A�`���E���͂��߂Ƃ��鍩���̂ق��ɁA�G�r�A�T���V���E�E�I�A�W�����A�E�~�K���A�t�N���E�Ȃǂ������悤�ɂȂ��
�����B���̊ԂɁuDNA�̃p�Y���v�����ׂ����A��Փx�𑝂��Ă���Ɗ����Ă��܂��B�����Ȃ��p�Y��������܂������A�u��炵�ג��ҁv�̎��̃X�e�b�v�A�b
�v��M���ē���ȃp�Y�����o�Ă���̂��y���݂ɂ��Ă��܂��B
|
���� �K��
�@
���{����w��w�@�@�������Ȋw������
�y����
|
2016.4.12
No.20�@����ʂ��Ė������݂�
�@����܂ł̃�
���[�R������ǂݕԂ��Ă݂�ƁA�u�����͒����ł͂Ȃ������v�ƌ�������Ă����������ق猩��ꂽ�̂����A���͂���̔�ł͂Ȃ����炢�����ł͂Ȃ���
���B����ǂ��납�A���I���`�ł������A�ہA���I���`�ł���B���߂ăI�I�X�J�V�o�������Ƃ��͑S�͂Ńn�`���ƐM�������A�A���W�S�N���E�X�o�J�Q���E�̗c����
�Ƃ������Ƃ�������ɂȂ��Ēm�����B�j�Ēp��������B��w�ŃV���r�A�V�W�~�������e�[�}�I�Ƃ����A���}�g�V�W�~�Ƃ̌�����������K���n���������B�M��
�̎ʐ^�i�ʐ^1�j������������肢��������Ǝv�����A�ߒ��Ԃ̎��������炵�āA�ǂ����Ԉ���Ă���炵���B
�@
�@�������A����Ȏ��ł��A�c�������璎���ώ@����̂͑�D���������B�Ȃ��ł��W�I�ɂȂ����̂́A�Ƃ̒��ɓ����Ă��钎�ł���B���Ƃ́A�_�˂̘Z�b�R�n�̘[
�ɂ���A�����ʂ�R�ɖʂ��Ă����̂ŁA���C��̓V��ɂ͂��������I�I�Q�W���������Ă������A�^�~�ɂ͌������ŃU�g�E���V���g���Ƃ�悤�Ȋ��������B����
�́A�߂܂��������ώ@����̂ɁA�C���X�^���g�R�[�q�[�̋�r�����p���Ă��āA�ꎞ�A�����̉a���ɗN�����C�����V�i�����炭�m�V���}�_�����C�K�j�ɐS�����A
�r�ɃA���ƃC�����V�����W�����āA���̍s�����������̂����ۂɂȂ��Ă����B�C�����V�́A�₪�Đ����ɂȂ�A��������A�܂��V�����C�����V���a������B
�������o���Ă��r�̒��ł͐��Ǝ����J��Ԃ���A���ɂ͂��ꂪ�i�v�@�ւ̂悤�Ɏv���A�ƂĂ��Ȃ��킭�킭�������̂��������A�A�������ׂč����ɂȂ������A
�C�����V�����̓p�^���Ƒ�����������Ă��܂����B�܂��A���鎞�́A�z�~���̃N�T�M�J�����V�����\�C���W�߂āu�L�r�v�Ə̂��A�Ƃ�ł��Ȃ�����i���C�H�j��
����Ă��܂����A�ƕr������Ȃ���j���j�����Ă����̂����̊ԁA�Ă̒�A���ł����Ă��܂������Ƃ�����B����ɁA�z�c�ɕ��ꂱ���J�f��߂܂��ĕr��
����A��ō̂��Ă����A����`���E�Ȃǂ�^���ẮA���J�f���H�炢���l�������J���قnj��߂�̂��D���������B
�@
�@�����āA�����A�������߂Ă���B�~�c�o�`�ƁA����Ɋ���A�J�����_�j�i�ʐ^2�j�ł���B���̃_�j�́A�̒�0.1mm�Ɣ��ɏ������A�~�c�o�`�̋C
�ǂ̒��ŔɐB���C�ǂ��ǂ��̂ŁA�~�c�o�`�͔��M�ł����ɁA����ł��܂��B���{�ɂ́A�×���萶������j�z���~�c�o�`�i�ʐ^3�j�ƊC�O���瓱�������Z�C���E
�~�c�o�`�i�ʐ^4�j�����邪�A�A�J�����_�j�́A�j�z���~�c�o�`�ւ̔�Q�������Ȃ̂ɑ��āA�Z�C���E�~�c�o�`�ւ̉e���͂قƂ�ǂȂ��B���̈Ⴂ�͂Ȃ�����
�������ƒ��ׂĂ��������ɁA�ǂ����A�~�c�o�`�̃O���[�~���O�s�����W���Ă��邱�Ƃ��킩���Ă����B�A�J�����_�j�́A�V�����h��Ɉړ�����ۂɁA��U�C
�ǂ���o�ă~�c�o�`�̑̕\������̂����A�Z�C���E�~�c�o�`�͑̕\�ł����߂��_�j�����m���A��������Ƃ����Ƃ��ł���̂ɑ��A�j�z���~�c�o�`�͓݊���
�قƂ�NjC�Â����A���Ƃ��C�t���ĕ������Ƃ����Ƃ��Ă����܂������Ȃ��̂ł���B���́A���̌��_�ɂ�����܂łɁA�~�c�o�`�ƃ_�j���Ђ�����ώ@���������B
����́A�k�}�̐�ɂ܂т������������̃u���V�ŁA0.1mm�̃_�j���������グ�A�~�c�o�`�̔w���ɂ̂��āA�~�c�o�`���ǂ̂悤�Ȕ��������邩�A���x
�O���\�~���O���s�����A�_�j�����Ƃ����Ƃ��ł��邩�ׂ�Ƃ������̂ŁA���ꂱ�����C�߂��̃f�[�^���L�^�����B�����Ɗώ@�𑱂��Ă���ƁA�����
������Ɗᐸ��J�ɔY�܂����̂����A�~�c�o�`���ꐶ�����w����~���p���ƂĂ����炵���A�������̂ł���B�܂��A�Ƃ��ɂ������ɂ������ƒ������߂�
������Ƃ������Ƃ��y�����Ďd���Ȃ������B���̎��ɁA�ӂƁA�`���ɏ������悤�Ȑ̂̋L������݂������Ă����Ƃ����킯�ł���B
�@
�@�����A�Ȃ��~�c�o�`�ƃA�J�����_�j�̊W�ɒ��ڂ������Ƃ����ƁA�����ɁA�l�Ԋ����Ɛ������l���̖�肪�W��Ă���Ɗ���������ł���B�A�J�����_�j
�̈�`�q�ׂ��Ƃ���A���̃_�j�̓Z�C���E�~�c�o�`�̓����ƂƂ��ɁA�C�O�������Ă����\�����������Ƃ��킩�����B�j�z���~�c�o�`�́A�܂��A�J�����_
�j�ɑR����p���������킹�Ă��炸�A���̃_�j�̐N���ɂ���āA�����������Ԃ܂�Ă���B�܂�A�l�Ԃ���������Ȃ���A�j�z���~�c�o�`�ƃA�J��
���_�j�͏o���Ȃ������͂��ŁA�j�z���~�c�o�`�ɂƂ��Ă͍~���Ă킢���Г�Ƃ����Ƃ��낾�낤�B����ɁA�j�z���~�c�o�`�͑��푽�l�ȍݗ��A���̎Ɋ�
����Ă��邽�߁A�j�z���~�c�o�`�̐��ނ����Ԍn�̃o�����X��傫���h�邪�����Ƃ��z�肳���B����Ɠ����ɁA�ߔN�A�l�I�j�R�`�m�C�h�n�_�n�`�ւ̎��S
�e���݂̂Ȃ炸�A�Ɖu�ቺ��s���ُ�̉\�����w�E����Ă��邱�Ƃ��ӂ݂�ƁA�j�z���~�c�o�`�ƃA�J�����_�j�̊W�ɔ_���W�ł���Ƃ͂ƂĂ��v��
���A��������ډ��������̂Ƃ���ł���B���̂悤�Ȑl�Ԋ����ɂ���Đ��������Ԍn�̘c�݂͂Ƃǂ܂邱�Ƃ�m�炸�A��̓I�ɂǂ̂悤�ȑ���Ƃ�ׂ��Ȃ�
���A��X�͍��A�����ɔ����Ă���B�����āA�������ڂ��Ă���~�c�o�`�ƃ_�j�Ɣ_��̊W�𖾂炩�ɂ��邱�ƂŁA���̓����Ɉ���߂Â���ƐM���Ă���B
�@���ł��A�������߂鎞�A�r�̒���`�������̂悤�ȍ��g���������邪�A�����ƈ���āA���̐�ɉ�X�̖���������A�Ƃ����ӎ�������B�����������҂́A��
�l�̎�Ƃ������A�l�̋����̕����܂܂ɁA�D���Ȃ��̂�ǂ��Ă��܂������ł���B�������A�����ے肷�����͂Ȃ��B���ꂪ�l�Ԃ̖{�\�ł��萶����
���ł���B�������A�����猤��������������Ă��鎄�̏ꍇ�A�����̌����ɂǂ������Ӌ`������A�ǂ̂悤�Ȗʔ���������̂��A�������ɂ͖��ɗ����Ȃ��Ă���
���I�ɂǂ����������Ƃɖ𗧂\��������̂��A�Ƃ������Ƃ������ɐ�������`��������B�D���Ȃ��Ƃ�������肽���Ƃ����̂Ȃ�A����ł���Ă��������A
�ƌ����Ă��d���Ȃ��B���E�ɏA���Ă���́A����n�������̂̒S�����A�{�I�ƁA��ʎs���̕��X���炲�v�]�₲�ӌ������������@��p�ɂɂ��邽�߁A�����
�݂𐳂����Ƃ������Ȃ����B�����A����Ȍ��ꂵ���b�͔����ɂ��Ă��A���łɃ~�c�o�`�̈��炵���ɂ̓��������Ȃ킯�ŁA���ꂩ������������������B���̐��
�������B

�ʐ^2�@0.5mm�V���[�v�y���V���c��̃A�J�����_�j�i�B�e�F��{���q�E�O�c���Y���j�D�����痑�C�c���C���X�����C�I�X�����D

�ʐ^3�@�j�z���~�c�o�`

�ʐ^4�@�Z�C���E�~�c�o�`�i�B�e�F��l�E�����M���j
|

�ʐ^1�@�ʼn_�ɕߒ��Ԃ�U��M��
��{�@���q
�������������E������
|
2016.3.2
No.19�@��l�Ƃ̎v���o 2013 : ��Ɗx�ŋN�������ƁB�@
�@�A���̎�q��
��̂��痣��Ĉړ����邱�Ƃ���q�U�z�Ƃ����D�Œ����Đ������闤��A���ɂƂ��āC����ꏊ���痣��āC�����Ɉړ��ł���̂́C���̎�q�U�z�̒i�K�̂�
���D��q�U�z�̗l���͑��l�ŁC�d�́C���C���Ȃǂ̔���}�̂Ƃ�����@�����łȂ��C�����Ɏ�q���^��ł��炤�Ƃ������@������D���̒��ł��C��q���A��
�ɉ^��ł�����Ă���A���i�A���U�z�A���j�͂��Ȃ葽���C�X�~����J�^�N���Ȃǐ��E��90��2,800��قǂ��m���Ă���D�A���U�z�A���̎�q�̕\�ʂ�
�́C�A�����Ăъ鎉���ɕx�G���C�I�\�[���ƌĂ�鏬���ȕt�������t���Ă���D�A���͂���ɗU������C��q���^������̂��D
�@���{�̒�R��ł́C�X�~����J�^�N���ȂǕ����̎�ŁC�A���ɂ���q�U�z���m�F����Ă��邪�C�X�ь��E���鍂�R�тŃA���U�z�A���̒��������������͋ɂ߂ď��Ȃ��D�u���R�A���̏����v�ƌĂ��R�}�N�T Dicentra peregrina�i�R
�}�N�T���C�P�V�ȁj�̎�q�ɂ̓G���C�I�\�[�����t���Ă��邱�Ƃ��m���Ă���i�}1�j�C�A���U�z�A���ł��邱�Ƃ���t�E�����i2006�j�ɂ���Ďw�E����
�Ă������C���ۂǂ̃A���킪�R�}�N�T�̎�q���^�����邩�Ƃ�����O�ώ@�͂���܂ŕ���Ă��Ȃ������D�Ȃ��C�R�}�N�T�͖k�C���Ɩ{�B�̍��R�тɕ��z����
���N���̐A�����D�Ԋ���5�`8�����ŁC4�����Ԃ��炾��ƉԂ��炭�i�}2�j�D
�@2012�N�C���͐M�B��w�Ƀ|�X�h�N�Ƃ��ď������Ă���C�k�A���v�X�ŎR�x���A���ނ̐��ԂƑ��l�����������Ă����D���ɏ�Ɗx�ŏW���I�ɒ�����W�J����
�����D��Ɗx�͖k�A���v�X�̓�[�Ɉʒu���C������i�W��3,026m�j�����Ƃ���R�X�̑��̂ł���D��Ɗx�͎R���t�߂܂Ŏ����ԓ��i��ƃG�R�[���C���j
����������Ă���C�o�X��^�N�V�[�𗘗p����C���{�w����3���Ԓ��x�ŎR���t�߂܂ŃA�N�Z�X�ł���D
�@����܂ł̒�������C��Ɗx�k���̕����E��Ɠo�R�������̊ێR�t�߁i�W��2,510���j�ɐ��S���̃R�}�N�T�������鋐��ȌQ��������i�}3�j�C���̌Q���̎��͂ɂ̓^�J�l�N�����}�A��Formica gagatoides �i���}�A�����C���}�A�����ȁj�����z���邱�Ƃ�˂��~�߂Ă����D�����E��Ɠo�R����2005�N�x�ɐV�݂��ꂽ�o�R���ŁC�A�N�Z�X�������l�ʂ肪���Ȃ��D�����n�Ƃ��Ă͂����Ă��ł���D
�@�^�J�l�N�����}�A���͖k�C���Ɩ{�B�̍��R�т̋�Ւn�ɉc�����C��������5�`9�����{�ł���D�^�J�l�N�����}�A�������{�ɕ��z���邱�Ƃ��������ꂽ�̂�
1970�N��㔼�ł���C���ł����̐��Ԃ͂قƂ�Ǖ������Ă��Ȃ��D���z��Ɛ��������R�}�N�T�Əd�����Ă���C�R�}�N�T�̎�q���^������A���̑�P���
�ł���D�����ŁC���͖�O�ō̎悵���R�}�N�T�̎�q���^�J�l�N�����}�A���ɒ���Ƃ����P���Ȏ������l�����D
�@2013�N��5�����{�C���͖�O�����������Ȃ����߂Ɋ��ȂƗі쒡�ɃR�}�N�T�̎�q�̍̏W����\�������D��Ɗx�̎R�����͍��������̓��ʕی��Ɏw
�肳��Ă���C���ׂĂ̐A���E�����̍̏W�ɋ����K�v���D�O�N�����\�����o����1�������x�ŋ��������Ă����̂ŁC�u���N��7�����ɂ͒������X�^�[�g
�ł��邾�낤�v�ƍ��������Ă����D�������C���ۂɑS�Ă̋�������ꂽ�̂�8���̉��{�ł������D�������C�̏W�ł���R�}�N�T�̎�q�̐��͐\������1/10
�ɍ팸����Ă����̂ł���D���̂Ƃ��͏ł����D8�����{�̏�Ɗx�̎R���Ƃ����Ώ��Ⴊ�~���Ă����������Ȃ������̂����C���V�����C��Ƃɍs�����Ƃ͏o
���Ȃ������D�������C9��13�`16���͓��{�����w��D�y���ł������D���ی��n�ɂ��ǂ蒅�����̂�9��20���ɂȂ��Ă��܂����D
�@�ߑO8���C���͖��F�ł��鏬���M���Ƌ��ɐM�B��w���{�L�����p�X���o�����C��Ɗx�Ɍ��������D��O�����ɍs���ɂ͒x���o���̂悤�Ɏv���邪�C���͂��傤
�ǂ����D�����s���Ă��H�ʂ��������Ă��āC�Q�[�g���J���Ȃ�����ł���D9�����{�̏�Ɗx�͂��łɏH��ʂ�߂��C���~���D�������C���̓��͍ō��̓V��
���C�_�ЂƂȂ��D�V�ŁC�����Ȃ��C10�����߂���Ƃ���C�����オ�����D
�@11���C�R�}�N�T�̌Q���ɓ���������X���^�J�l�N�����}�A���͊��}���Ă��ꂽ�D�Ȃ�ƌ�����s�̌���ɂ�����o���킵���̂ł���D�����n�̎��͂�V��
���ƗY�A������щ��C�n�ʂɂ͒E���������������C������ꂽ�i�}4�j�D�A���w�҂Ƃ��Ă͋����̋ɂ݂��D���E�Ŏn�߂ē��{�Y�̃^�J�l�N�����}�A���̌�
����s���m�F�����̂ł���iUeda & Komatsu
2014�j�D�������߂��ʂȂ�C��X�̓R�}�N�T�̎�q��T���n�߂��D���ׂẴR�}�N�T�̉Ԃ͌͂ꗎ���Ă���C�Ȃ��Ȃ���q�͔����ł��Ȃ��D���炭�̒T
���̌�C�͂ꂽ�Ԃ��܂��s�ɕt���Ă���R�}�N�T�������������ł����D�Ԃ̒��ɂ͎�q�������Ă���C�v10�̎�q���m�ۂ��邱�Ƃ��ł����D
�@�Q�̃A���̑��̓�����̂������Ɏ�q��5���u���C�A���̎�q�ɑ���s�����P���Ԋώ@�����D�A���̎�q�ɑ���s���́C�@������o�Ă���������
�����A������q�ɋ��������߂��C�A��q�\�ʂ�G�p�Ń^�b�s���O����(�}5)�C�B�G���C�I�\�[���̕������{�ł��킦�C�����ɉ^�э���(�}6)�ł���D�P��
�Ԃ�10�̂���5�̎�q�������ɉ^�э��܂ꂽ�iKomatsu et al. 2014�j�D
�@�a���������Ȃ����R�n�тɂ����āC���b�_�𑽂��܂ރG���C�I�\�[���̓A���ɂƂ��Ė��͓I�ȉa�����ƂȂ�D�^�J�l�N�����}�A���̑����ɉ^�э��܂ꂽ�R�}�N
�T��q�́C�G���C�I�\�[���݂̂��H�ׂ��C�s�v�Ȏ�q�{�̂͑��O�Ɏ̂Ă���̂ł��낤�D�̂Ă�ꂽ�ꏊ�ŃR�}�N�T���V���ɉ萁���C���������z���L����
����\��������D�{�����̐��ʂ́C��ł��뜜����鍂�R�A���ƍ����Ԃ̑��ݍ�p�𖾂炩�ɂ������̂ł���C���R���Ԍn�̕ۑS�헪�ɑ��Q�W�S�̂̕ۑS��
������̈Ă���邾�낤�D
�@���̓���7���Ԃ̒����ŁC��X�͈ȉ��Q�{�̘_�����o�ł����D
Komatsu T, Itino T, Ueda S (2015) First report of seed dispersal by
ants in Dicentra peregrina (Papaveraceae), an alpine plant in the
Japanese Alps, Entomological Science, 18(2): 271-273
Ueda S and Komatsu T (2014) Observation of nuptial flights of an alpine
wood ant, Formica gagatoides Ruzsky, 1904 (Formicinae) in the Japan
Alps, ARI Journal of the Myrmecological Society of Japan, 36: 7-10
�@7���Ԃ̖�O�����Ř_��2�{�Ƃ����͎̂��ȐV�L�^�ł���D���ꂪ�Ȃ��������R�́C�\�������ōD�K�Ȓ����n��ݒ肵�C�������������Ă������ƁC���{�̍�
�R�тƂ����N�������������Ȃ��ꏊ�Ō����������Ȃ������Ƃ���������D�����M���Ƃ�����L�ȋ��������҂Ɍb�܂ꂽ���Ƃ��傫���D
�@���{�̍��R��͂܂��܂��������s���͂��Ă��Ȃ��t�����e�B�A���D�{�R���ΏۂƂ�����O�����͐V�K���������p�C�I�j�A���[�N�ł���C�l�X�̍D��S���h������D���ɁC�A���ƍ����Ԃ̎�ԊW�ɂ��Ă͖��𖾂ȕ����������C���㑽���̐V�m���������邾�낤�D
<�t�}����>

�}�P�D�R�}�N�T�̎�q.��q�{�̂͌����鍕�F,�G���C�I�\�[���͖ȏ�Ŕ��F�i�B�e�F�����M�j

�}�Q�D��Ɗx�R�����̍��I�n�ɍ炭�R�}�N�T.���̔��������獂�R�A���̏����ƌĂ��
�i�B�e�F��c�����j

�}�R�D��q���̎悵���R�}�N�T�D�Ԃ͌͂�Ă��邪�C�܂��s�ɕt���Ă����i�B�e�F��c�����j
 �D �D
�}�S�D�E�������^�J�l�N�����}�A���̏����i�B�e�F��c�����j

�}�T�D�R�}�N�T�̎�q�\�ʂ�G�p�Ń^�b�s���O����^�J�l�N�����}�A���̓����A���i�B�e�F�����M�j.

�}�U�D�^�b�s���O��,�G���C�I�\�[���̕������{�ł��킦,�����Ɏ����A�����i�B�e�F�����M�j�D
|

��c ����
���{����w�E�������Ȋw��
|
2015.11.24
No.18�@�T���������ɁF�w���ƃ{�[�g�Ǝ��X���P�b�g�@�哇�@�ꐳ
�@10���̉Ηj���̓��P�b�g�̑ł��グ�����邩��C���C�g�g���b�v�͊C�̋߂��̃T�o���i�ł�낤�D�ł��グ������̂ɂ��傤�ǂ����r�[�`������炵������C���C�g���Z�b�g������r�[�����ĕl�ӂɍs�����I
�@�p�� Paris �̃I�����[ Orly
��`�ŁC�����͋C�܂���ȃt�����X�̋��������҂���������̓I�Ȓ����v������b���Ă����D���������̑��̃X�P�W���[���́C�Ƃ���t�B�[���h�X�e�[�V����
�i���{�Ō����Ƃ���̉��K�т̂悤�ȂƂ���j��2�T�ԑ؍݂���ȊO�C�����ʂ���ɉ������܂��Ă��Ȃ��炵���D����ȉ�b�����Ȃ���C�t�����`�M�A�i
(�t�����X�̃M�A�i) �s�� AirFrance �ւ̎�ו��J�E���^�[�̒�����ɕ��ԁD
�@�Ƃ肠�����C���n�ɓ���������C���ꂼ��̌����҂����ꂼ��̕��ތQ�ɉ��������@�ŁC�t�B�[���h�X�e�[�V�����̃g���C��������������C���ɂ͐[���W��
���O���̒��ɓ������肵�ĉ�������̖ʔ������Ɓi���j��T���D���ꂪ�����̃t�����`�M�A�i�����ɂ�����~�b�V�����ł���D��t�����̖��т�����������
�̏W�ɍœK�Ȃ킯�ł͂Ȃ��C���ɂ̓A�X�t�@���g�ܑ����ꂽ���H�̗��e��w���|�[�g�̂悤�Ȃ�����x�̏�����ꏊ���ʔ������Ƃ�����D�Ƃ肠�����́C
�������Ȃ����Ƃɂ͎n�܂�Ȃ��D�����Ė�͖��Ӄ��C�g�g���b�v������D
�@���āC�w���|�[�g�C�Ƃ������t���o�Ă������C�ʐς̑啔���i90���ȏ�j���ˑR�W�����O���ɕ���ꂽ�t�����`�M�A�i�ɂ����āC�w���R�v�^�[�͂ƂĂ��d�v
�Ȉړ���i�ł���i�ʐ^1�j�D�����v�揑�̌o��̍��ڂɂ́C�w���R�v�^�[�Ɋւ����p���p�ɂɓo�ꂵ�C��̂̃t�B�[���h�X�e�[�V�����̓w���|�[�g�������
����D�w���ɏ��C�Ƃ����̂́C�ŏ��ɗ������͋��������C�����Ɋ���Ă��܂����̂ŁC�u���̑O�o�����R�̂Ă��J���Ă�������C���̒����̎��̓w����
���d�@���^��ŎR���Ń��C�g�g���b�v�����悤�v�ȂǂƂ����A�C�f�A�����ʂɏo�Ă���悤�ɂȂ��Ă��܂��D
�@�w���ƂƂ��Ƀt�����`�M�A�i�̃W�����O���ŏd�v�Ȉړ���i�̓��[�^�[�{�[�g���D���������[�g���قǂ̃{�[�g�ɉו��ƂƂ��ɏ�荞��ŁC�Ђ�����㗬���
�w���D�p������̒��s�ւ��������錧�����ݒn�i�t�����`�M�A�i�̓t�����X�̊C�O���j�̃J�C�G���k Cayenne
����C�Ԃ�2���Ԃقǂ����đ傫�Ȑ�ɍ��ꂽ�����Ȕg�~��ɒH�蒅���C�������炳��ɐ����Ԃ����Đ��k�シ��i�ʐ^2�j�D���͉��V���̊��G�ɂ����o��
���Ȃ����C���ʂ�����������͂Ƃ���ǂ��됅���Ԃ����オ��}���ƂȂ��Ă���C���������ꏊ�͑D���̘r�̌������炵���D���V���ł̐����Ԃ̑D���͂���Ȃ�
�ɔ��邪�C���ʂ������Đ��ʂ������o������ɍ炭�J���S�P�\�E�̉Ԃ�C���X�����ʉ߂��Ă��������t�H�`���E�C���s�����Y��ȃc�o���K�Ȃǂ̂�������
�S���O���邱�Ƃ͂Ȃ��D
�@���āC����Ȉړ���i�����ł��\���h���I�ȃt�����`�M�A�i�����Ȃ̂����C�����Ō����鍩������������炸��X���y���܂��Ă����i�Ƃ������C�����炪
��ړI�Ȃ̂ł����łȂ��ƍ���j�D���̐��͗����ڂł���C���Ƀz�\�K�ȂƂ��������ȉ�ނɏœ_�ĂČ��������Ă���D���̃O���[�v�͂�����u�G����
���v�ł���C�c�������t�ɐ����ĉ߂����T�^�I�ȃ��[�t�}�C�i�[ leaf miner �ł���D
�@�����ĉ�X�����ڂ��Ă���̂́C�c�����̑O���ʂ̃��[�t�}�C�i�[�Ƃ��ĉ߂����C�c�����̓r������S�[�� gall (���)
�ƌĂ��J���X��̑g�D��A���̂Ɍ`������z�\�K�ł���i�ʐ^3�j�D���́u�r������S�[�����`������v�Ƃ����̂��~�\�ŁC�{���ɂ���܂ł͂����̃��[�t
�}�C�i�[�Ȃ̂ł���D��Ɍ��܂����������S�[���`�����˔@�Ƃ��Ďn�܂�悤���D���[�t�}�C�i�[�Ƃ��ĉ߂����Ō�̗���ƁC�S�[���`�����n�߂����Ƃ̊�
�ŁC�����������ʂ��قȂ��`�q�Ƃ��͂Ȃ��̂��낤���H
�@����Ȏ��������̎���Ȃ�\�ł���D�Ƃ������C���悤�Ǝv���Βn���̗����̃W�����O���ł��C�y�ɗ���Ă��܂��D�\�Z�̐\���Ɗl�����܂߁C����Ȃ�ɋ�
�J�͂��邪�C���Ȃ��Ƃ���s�@�͖������ł��邵�C�Q�m�����K�͉�͂��Ă����@�B�����݂���D�{���ɂ�������Ɍ������ł��Ă���Ȃ��Ǝv���D
�@�����č��C�܂��Ƀt�����`�M�A�i�ɂĂ��̌��e�������Ă���̂����C����̒����ŐV���ȁu�r������S�[�����`������z�\�K�v�����������D���������̎��
����܂ʼn�X�����ڂ��Ă���������̐��������C�������J�C�G���k����Ԃ�1���Ԃقǂ̃N�[���[ Kourou
�Ƃ������̋߂��ł���������D�w�����{�[�g���K�v�Ȃ��D���䌳�Â��Ƃ͂��̂��Ƃ��D�Ƃ����킯�ŁC���͂��̎��ݑ㎔�炵�Ă݂�v���W�F�N�g�𗧂�
�グ�悤�C�Ƃ������ƂɂȂ����D
�@���āC�b�̓��P�b�g�ɖ߂�̂����C�N�[���[�ɂ̓t�����X�̃��P�b�g�ł��グ�Z���^�[������D������Ɠ����ɓ������Ƃ���ɂ́C���V�A�̃\���[�Y�̑ł���
���Z���^�[������D���Ƃ������C�F����g�߂Ɋ�����ꏊ�Ȃ̂ł���D���C�g�g���b�v�̃Z�b�g�A�b�v��C�N�[���[�̎s�X�n�Ńr�[�������݁C�߂��̕l�ӂ�
�Ƌ}���D���łɑ����̐l���l�ӂ̊��ɍ��|���Ă���C�݂�Ȃ��ł��グ�Z���^�[�̂��鐼�̕��p�����Ă���D�ł��グ�\�莞���͌ߌ�6��34���ł���C����
���Ǘ[���������ギ�炢���D
�@����Ȃ���ȂŁC�吼�m�̕l�ӂɂāC��n�ɒ��ޑ��z�����Ȃ���r�[���Ŋ��t�D���ꂾ���ł��������߂��ˁC�Ƃ��݂�ȂŘb���Ă�����C�˔@���̕��p������
���Ȃ����i�ʐ^4�j�D������������ł��������z��������x�����Ă������̂悤�ȃI�����W�F�̌��̉���C�������ƁC�ł��m���ɓo���Ă����D�܂�Œ�
��������̂���Ō��Ă��邩�̂悤���D�Ȃ�قǁC�V�ˉ��`�����̉̂́C�����������Ƃ��̂��Ă����̂��C�Ǝv�����͓̂��{�l�̎��������낤���ǁC������
�����傤���Ȃ����Ƃ��l���Ă��邤���ɂ����P�b�g�͂ǂ�ǂ�쓌�ւƐi��ōs���D
�@���P�b�g�̔������͂������ɂ́C���̏�ɔ�s�@�_�Ȃ�ʃ��P�b�g�_���i�ʐ^5�j�D�܂�Ń}�C���̂悤���C�ƌ����҈ꓯ�吷��オ��D�����āC����̊ϏO���ł��グ�̐������j���đ傫�Ȕ�������Ă���D�܂�ʼn����̂��Ղ�̂悤���D
�@���P�b�g�̑ł��グ�Ȃ�āC�����ɏI���C�x���g���낤�Ǝv���Ă������ǁC�V��ɂ��b�܂�C2�̃u�[�X�^�[���藣����ė������Ă����܂ŁC����3��
���炢�͌��Ă����Ǝv���D�����āC���ꂾ���������ԃ��P�b�g�����Ă���ƁC�F�X�Ȃ��Ƃ��v���o���D���������Ώ��������C�e���r�Ń��P�b�g�̑ł��グ������
�̂��D���ŁC�����͎��ۂɌ��Ă݂����Ǝv���Ă������ƁC����ɂ́C�����͐����w�̓��ɐi�����Ǝv���Ă������ǁC���Z�ŕ������K���Č��\�{�C�ŕ����w����
�������Ǝv�������ƂȂǁD
�@���Z�ŏK���������́C�����������P�b�g�̑ł��グ�����Ċ��������N���N���̐��̂�\���Ă��ꂽ�C������D�܂�C����ȋ�����̂��ǂ̂悤�Ȏd�g�݂ʼnF
���܂Ŕ��ł����̂��C�����āC�F���̉ʂĂ͂ǂ��Ȃ��Ă���̂��H�@�܂�C�����͗������ۂ����Ƃ��D���Ȃ낤�ȂƎv���D�����琶���w�̓��ɐi���
���C�핪���Ƃ����K���̈�`��ՂƂ��C���������e�[�}���D���Ȃ낤�Ȃ��C�Ɩ��Ƀ��P�b�g�����Ȃ���[�����Ă��܂����D����ȕ��ɂ��āC�핪���Ɗ��
�]���ƉF���̉ʂĂɎv�������炵�Ȃ���C���߂ă��P�b�g�_�̃}�C���߂Ă݂�ƁC�}�C���̐�[�ɂ�����̉S�[���Ɍ����Ă���D
�@�S�[�����`���ł���悤�ɂȂ邽�߂ɂ͂ǂ̂悤�Ȉ�`�I�ύX���K�v�������̂��낤���C�����āC���[�t�}�C�i�[�Ƃ��ėc�������������鍩������������
���ŁC�ǂ����č������悤�ȃz�\�K�͗c�����̓r������S�[�����`������悤�ɂȂ����̂��H�@����ȁC���ߗv���Ƌ��ɗv���Ɋւ���^�₪�ǂ�ǂ�N��
�Ă���D����ς藝�����ۂ��b�ɂȂ�Ȃ��C�Ǝ����ł��v���ƂƂ��ɁC���������Ȃ������獡��̃��P�b�g�̑ł��グ�����邱�Ƃ͂ł��Ȃ������̂��낤��
���C�Ƃ��v���D�s�v�c�Ȃ��̂ł���D
�@���āC����Ȋ����Ńt�����`�M�A�i�ł̒����͂����\�z�O�̎h����^���Ă����D�������ɂȂ邩�͂킩��Ȃ����C����͂ǂ�ȋ������҂��Ă���̂���
�����D�\���[�Y�̑ł��グ�ł���������ǁC�ł���u�r������S�[�����`������z�\�K�v�̌�������C�傫�ȋ�����������Ǝv���D�t�����`�M�A�i�̃z�\
�K�������C���낻��P�Ȃ� expedition ���� experimental biology
�ւƁC����̃��P�b�g�̗l�ɂ��܂��O���ɂ̂��Ă����������̂ł���D

�ʐ^1. �w���R�v�^�[����̃A�}�]���M�щJ�т̒��߁D

�ʐ^2. �{�[�g�ɏ���Đ������Ă���l�q�D���݂Ƃ��ɁC������̔M�щJ�тł���D

�ʐ^3. �c�����̓r������˔@�Ƃ��ăS�[���`�����n�߂�z�\�K�D�Ԃ��Ȃ��Ă��镔�����S�[����̕����D���A���̓t�N�M�Ȃ� Clusia blattophila.

�ʐ^4. ���P�b�g�����܂��ɋ�֏オ���Ă������Ƃ��Ă���u�ԁD�[���Ɠ������炢���C����ȏ��ῂ����D

�ʐ^5. �}�C���̗l�Ɍ����郍�P�b�g�_�D�E���ɂ́C�藣����ė������Ă���2�̃u�[�X�^�[��������D |
�哇�@�ꐳ
�@
���s�{����w��w�@�������Ȋw������
����
|
2015.04.20
No.17 �鋫 ���V����ڎw���� �@��
�@1998�N8���B��w2�N��
���������́A�V���̓V�C�}�Ƃɂ�߂���������������Ă����B���̔N�A�����{�̊e�n�Ō������Ă�������n��̃I�I�M�������}���A�O���̖k��ƂƂ��ɕx
�R���ɂ���Ă���̂����������Ƒ҂��\���Ă�������ł���B���̔N��8���ɂȂ��Ă��O�����{�B�ɒ���Ă��ĉJ�����������B1998�N8��24���̒��A�V
���̓V�C�}������ƁA�O�����r��Ėk���n���ɓ삩��̕������ꂱ��ł����B�����͑_���ڂ��B�ċx�݂Ŏ��ƂɋA�Ȃ��Ă������́A�I�I�M�������}��������
�Ȃ炱�����낤�ƁA8���ɂȂ��Ă���2����1�x�̃y�[�X�Œʂ��l�߂Ă����C�݉����̒r�ɕ��Ɠ�l�ŏo�������B�ڂɂ���̂̓M�������}����B���������ҊO
�ꂩ�Ǝv���ċA�낤�Ƃ����Ƃ��A�r���Q���ԑ����̃M�������}�ɍ������āA��⍕���̋��������}��1���������Ă��邱�ƂɋC�������B�������畃�Ɠ�l
�ŁA�������ł��̃g���{��_�����B30���قǂ����āA�悤�₭�̏W���Ă݂�ƁA�܂�����Ȃ��x�R�����L�^�̃I�I�M�������}�������B�̏W��������ɁA�߂�
�̍H������̊ē��̐l���琺��������ꂽ�B�������Ă���̂��������Ǝv���Ă������A��l�����܂�ɂ��^���Ȃ̂Ő����������Ȃ������̂��������B���̓�
�̗[���A�x�R���̃g���{���ꏏ�ɒ��ׂĂ����r�؍�������ɓd�b����ƁA�u�V�C�}�����č����͉������Ǝv���Ă�����ł����ǁA�d���������čs���Ȃ��������
����v�Ə������������������B
�@2001�N7���B��͂薈���̂悤�ɓV�C�}�߂Ă����B�悤�₭�̏W�������肽�k�A���v�X�̉��n�A���V���֒����ɍs���������l���Ă�������ł���B
�ŒZ�̓o�R���R�[�X�^�C����12���Ԕ������鍂�V��������ɂ́A�Œ�ł�3���͕K�v�ł���B�����A���͑�w�@�̏C�m1�N���������B�����璲���ɍs����
�ƌ��߂����A�u�����̂���3���قNjx�݂܂��v�A�Ɠ����̎w�������Ƀ��[�������āA���̂܂ܓo�R���ւƋ}�����B�_���͍����ɐ������鍕���Ȃ�A�J�g���{��
���A���c�A�J�l�ł���B�k���n������͖��L�^�ŁA�x�R�����甭�������\���͒Ⴂ�Ə����ꂽ�������������̂����A�n�}�߂Ă��č��V���Ȃ炢�邩��
����Ȃ��ƑO����ڐ������Ă����̂ł���B�������A���V���ɓ������������ɑ������璲�����Ă����̂����A����Ɍ�����Ȃ��B�[���ɂȂ�A���s�҂���A
�u�����������V���ɗ����̂�����A�L���ȍ��V������ɓ��낤�v�ƒ�Ă��ꂽ�B����ǂ��A�ꕪ�ł������̎��Ԃ̐ɂ����������́A���̂��U����f��A�r����
�Ɠ�l�ŗ[�����Ђ�����g���{��T���Ă����B�������ɊÂ��Ȃ����ƒ��߂����Ă����Ƃ��A�r����́u���܂����I�v�Ƃ������ѐ����������Ă����B�r�����
�Ԃ�����ƁA����͕�����Ȃ��k���n�����L�^�̃��c�A�J�l�������Ă����̂������B���Ȃ݂ɁA�L���ȍ��V������ɂ́A�����ɓ��������Ƃ��Ȃ��B
�@���́A���S���������獩�����D���ŁA���Ƀg���{����ԍD���������B�����ɂ͂��낢��ȃ^�C�v�̐l�����邪�A���͒������g���{�̐V�Y�n�T���ɓ��ɗ͂��
��Ă����B�������A���R�Ȃ��猋�ʂ̏o�Ȃ������قƂ�ǂ���B���ʂ��o�Ȃ��������������ƁA������y�[�W�������Ă���������Ȃ��C������B����Љ��
2�̗�́A���̗\�z���قڑ_���ʂ�ɓI�����������Ȃ���ł���B�����Ɋւ��Ă��A������v���Ԃ��ƈ���ԈႦ��Ό����Ƃ��Ă����\����������������
�ł���B���̌�A���m�����擾���A�{�i�I�Ɍ������s���悤�ɂȂ������݁A�����̏W�ƌ����͎��Ă���ʂ������悤�Ɋ����Ă���B�u�_���ʂ�ɗǂ����ʂ��ł�
���Ƃ͖{���ɏ��Ȃ��B�v�u����ǒn���ȓw�͂𑱂��Ă���A���܂ɂ͏d�v�Ȍ��ʂ��o��B�v�u�������A���ӂ��Ă��Ȃ��Əd�v�Ȕ����ɋC�Â����ɏI���̂���
����Ȃ��B�v�ȂǂȂǁB���́A���č��V���̒������v�悵�Ă����Ƃ��̂悤�ɁA�V���������Ɋ��҂�c��܂��Ď����v����l���ẮA���҂ƈقȂ�f�[�^���o
�Ă��邱�Ƃ��������A�M�d�Ȍ��ʂ������Ƃ��Ȃ��悤�ɐS�|���Ă���B
|
���U�Ƀ��J�V�g���{�̗c���������s���l�q
�@��
�@
�Y�ƋZ�p������������C������
|
2015.02.15
No.16 �����ƐA�� ���J���@����
�@�ǂ������킯���A���̐S�t����
���炸���ƐA�����D���������̂ŁA�����̖��m�ȋL���͏��w�Z��N���̉ċx�݂̎��R�����ł��B�i�{�[�����ɖ�R�̊G��`���A���ꂼ��̍����̐���ꏊ�ɒ���
�z�u���܂����B1970�N�㏉���͐��Ԃ̒��Ő����𗝉�����Ƃ��������������̂ł��̉e���������̂��ȂƎv���܂��B�����ɏZ��ł����̂ŁA�I�I�����T�L��
�ǂ�����N�k�M�̖��������̂ł����A��������̃J�u�g���V��N���K�^�͂��炸�A�J�i�u������B���̂����A�F�B�̉Ƃɂ������H���A���ɐS��D���A����
�Ɏ����Ă��܂��B
�@��w�͐A���w�����ő�w�@�̌������͐A�����ɂ������̂ŁA�w���ō����i��搶�̖��Ғœ����w����u���āAR.M. Alexander����́uThe
Invertebrates�v��ǂ�Ŗ��Ғœ����̑��l���ɖ��f���ꂽ�̂ƁA�ΐ쓝�搶�̃[�~�Ŋ���t�Y�搶�́u�����̐����w�v�N�����ēǂ�Ŗʔ���
�ȂƎv�������炢�ŁA�����Ƃ̐[���������͂���܂���ł����B��w�̃T�[�N���͐����w������ŁA�����������̍�{���炭���c�ӏG�V����A�N���K�^����
�G�ؗ�����A���S�ʂ̐[�Õ��n����Ȃǒ����͂��������̂ł����A�����ɍs���Ă����[�~�X�V�W�~���̓i�K�T�L�V�_���h�L�ɖ��͂������Ă��܂����B
�@���ɋ����������������͉̂������Ċ���ɗ��Ă���ł��B���C���Ă��炭���āA�����̖ї��G�Y�搶�����{�ɗ��āu���J���N�A�`���E���Ȃ����v�Ƃ���
�Ȃ肨��������āA����s�y�b�g�}���������āA�Z�p�x�����̕S�X�R��q����Ɏ����������o���܂����B���̌�A���̎O�}�L���搶����W���u�`�̋@�������
�����A�f���Ă��b�����Ă��邤���ɁA�O�}�搶���Ȃ̑c���̍]���O�搶�̂���q����ł��邱�Ƃ��킩��A�܂��A�O�}�搶���N���}�`�X�̈��ƂŐH���Ƃ���
�ȏ�ɐA���ɐ��ʂ���Ă������Ƃ�����A���̌�A���X���w�����������Ă��܂��B�܂��A��㏟�炭����͂��ߑ����̂���q����ɂ��䐢�b�ɂȂ��Ă��܂��B��
���搶�́A���{�ŕ��s�I�ɐ����Ă����`���E��DNA��p�����n����̓O���[�v�����܂��������ł��������Ȃ����ƍl�����A����DNA�������ݗ���
��܂����B���̉�͒��ނɌ��炸�A�����S�ʂɂ܂�����A�w���w���̂��납�炢�낢�낲�������������Ă������q�i���w�̑��V�ȎO�搶��A���ƂŏW�c��`�w��
����͂�����������{�b��搶�̍����w�҂Ƃ��Ă̈�ʂ��͂��߁A�����̂��Ƃ��w���Ă��������ƂĂ��L�v�ł����B�n�ӈ�Y�搶�ɂ́A�M�t�`���E�̏W�ɘA��
�čs���Ă��������ăJ���A�I�C�̐i���ɋ���������������������Ă�����������A���̌�����낢��Ȏ��Ō䐢�b�ɂȂ�܂����B
�@�����ʂł́A����n�a�̈��`���a�����A���w�����̈��y�̒˒J�T�ꂭ��Ɠ��{�ł͂��߂ẴG�{�f�{�̃V���|�W�E�����u�����ƐA���̔����̈Ⴂ�v��
�����e�[�}�ŊJ�Â��A������A���Ɣ�r���邱�Ƃ̖ʔ����ɋC�Â��܂����B�����āA����̏�쒼�l����\�����ꂽ���������̓���̈�ŁA�n�i�J�}�L��
�ƃR�I���M����������Ă�����n�����搶�̔ǂɉ����Ă��������A�����̕��X�ƂT�N�ԓ��������O���[�v�ɏ������A�����̎h�����܂����B���̔lj�c�̎�
�ɁA��D���̑q�J�����������Ɏt���Ƌ��A�lj�c�̋x�ݎ��Ԃ��ɒ����ɗ�݁A���̗���ŁA�������u��`�v�ɓ����ƐA���̔����̈Ⴂ�ɂ���2002
�N����Q�N�ԃ����[�Βk�������Ă��������܂����B���̓��e�A�Ⴓ�y��A���ł��ʔ����_�������̂Ő}���قŒT���Ă݂ĉ������B���̍��A�ړ�����b����������
�ł����A�ړ��������Ȃ������Ȃ́u�ߏ��ɂ����R�������Ă��v�Ƃ������炵�������ɂ��A�ЂƎR�����āi�R�т͂��ǂ낭�قLj����ŋ����قƂ�ǂ������
���̂ł����߂ł��j�A���T�A���̏W�ō������ɂ����������W�߁A�N���K�^�u�[���������Ă��낢��Ȏ�ނ̃N���K�^����Ă܂����B�����āA�s����w�ɏW��
�u�`�Ɏf�����Ƃ��ɂ��낢��i���ɂ��Ă��f���������Ƃ̂������ΐ�Ǖ�搶�́u�����̒a�� :
��疜��ւ̐i���ƕ����i�����V���j�v�ɏo�Ă��鍩�����V�~����`���E�܂ō̂�܂���A�̌n�����č����̕������邱�Ƃ��ł��A�܂��܂��������N���܂�
���B���̍��A�p�v�A�L���C���N���K�^�̐F�ʑ��^�ɋ����������A���{�̔|�{���̒I�����g���Ĉ�l�Ő��S�C�̌�z�������n�߂܂����B�����A�����o������Ɛ��b
�ɍ���̂ƁA�p�シ��ɂ�ߐe��z�̉e�����Ђǂ��A�U��قnjp�サ���Ƃ���Œ��~���Ă��܂��܂����B���̑����͒�N��ɂ�낤�Ǝv���Ă��܂��B
�@�ї��搶�̌����͂ǂ�ǂ�i�W���A�O�}�搶�A��t�G�K����A���؍F�i������n�߂Ƃ����搶���̂����͂̂��ƃZ�Z���`���E�̌n���ɂ��Ă̘_���\�ł�
�܂����iTanikawa-Dodo et al.
2008�j�B���̂���A����Ń[�u���t�B�b�V���Ő������v���������Ă����^��O���N���q���̂��납��n�i�J�}�L���̌������������������ǁA�ǂ�������
�̂��Ƒ��k�ɗ��܂����B����Ȗʔ������̊w�U���ʌ������ɏo�����為�������ʂ�Ɨ�܂����Ƃ���A�����Ȑ\�����������グ�����̑�����܂����B����ɁA�k
��_�w���ɂ����哇�ꐳ�N���A�w�U���ʌ������Ƃ��ăN���~�z�\�K�̃z�X�g���[�X�]���̎d���������ł�肽���Ƃ����Ȃ�K�˂Ă��āA�������ɉ�X�̃Q�m���Z
�p�Ƒg�ݍ��킹��A�z�X�g���[�X�]���̌�����`�q���P���ł������������̂ŁA�w�U���ʌ������ւ̐\���𗹉������Ƃ���A����܂��̗p����܂����B�A����
�W�Ő\�����Ă����������ɂ����̂ł����A�����ʂ�͍̂�������A�������A�ɂ߂Č��I�����[�h���[�J�[�̓�l���A�����肾�����������ɉ����A��
�܂܂łƈ�������͋C�ňꓯ�A�����ɐ���オ�邱�ƂƂȂ�܂����B�哇�N�́A�S�X���������s���A�ї��搶�ƎO�}�搶�����S�ƂȂ��Đi�߂Ă����^�e�n
�`���E�̌n���̘_�����n���ƃz�X�g���[�X�]���̊ϓ_����ʔ����܂Ƃ߂Ă���܂����iOhshima et al.
2010�j�B�n�i�J�}�L���̋[�ԂƃN���~�z�\�K�̃z�X�g���[�X�]��������`�q�͂����������i�s���ł��̂ŁA���ʂ͂��y���݂ɁB�܂��A�哇�N�ƃz�X�g���[�X
�]���̌����ɂ��Đ܁X���낢��c�_���Ă��������ɁA�u�����K���`���̐i���v�̑�\��ł��邱�ƂɋC�Â��܂����B�����K���`���Ƃ́A��̕ψقł͓K���x
���������Ă��܂����A�������̕ψق��d�Ȃ�ƓK���x��������悤�Ɍ�����`���ł��B�Ⴆ�A�z�X�g���[�X�]���́A�c�����V�����z�X�g��H�ׂ���悤��
�Ȃ�ψقƎ��e���V�����z�X�g�ɎY������Ƃ����Q�̕ω����N�����Ă͂��߂Đi�����܂��B�c���Ǝ��e�̗����̌`�����i������A�������A�����z�X�g�ɓ]����
��悤�ɐi�����邱�Ƃ͋ɂ߂č���Ɏv���܂����A�����ł̓z�X�g���[�X�]���͕p�ɂɋN�����Ă���A�Ȃ�Ƃ��s�v�c�ł��B���ꂪ��̂��������ƂȂ��āA
�T�̍b���̑q�J������A�A�Q�n�̋[�Ԃ̓������F����Ɩx���搶�A�����̋����̐[�Â���A�J�C�R�̃z�X�g���[�X�]���̓��c�������ƐV�w�p�̈�𗧂��グ
�邱�Ƃ��ł��܂���
�ihttps://staff.aist.go.jp/t-fukatsu/SGJHome.html�j�B
�@���āA����Ȃ킯�ŁA���͂����ς�A�����S�Ɍ�����i�߂Ă���̂ł����ihttp//www.nibb.ac.jp/evodevo�Ghttp://www.nibb.ac.jp/plantdic/blog/; http//www.nibb.ac.jp/evodevo/tree/00_index.html�j�A
���������A���ɍ����Ɣ�r����Ȃ����Ă��܂��܂����B�A�������Ȃ���A�����ł͂���Ȃ��Ƃ͂�����̂��ȁA�����̉��ɂ�����̂��Ȃ��A�Ƃ����l����
���܂��B�����́A�]�ʈ��q�ɂ���ĂЂƂ����܂�̑g�D����扻���Ĕ������܂����A�A�����Ԃ��`������Ƃ��Ɏ����悤�Ȏd�g�݂��g���Ă��܂��B�܂��A���
���[�𑝂₵�Ă����ҒŌ`���́A�A���̕����g�D���J��Ԃ��t������ĐL�тĂ���l���Ǝ��Ă��܂��B����A�זE�����Z�v�^�[�̕ω��œ����ꏊ��ς��邱��
�ɂ���ĐV�����튯�A�Ⴆ�Ί{���`�������Ƃ����悤�Ȃ��Ƃ́A�זE�������Ȃ��A���ł͖����̂��Ȃƒ��炭�v���Ă��܂����B
�@�Ƃ��낪�A�N�n�߂ɖʔ������Ƃ�����܂����B�_�[�E�B�����r�[�O�����ōq�C�������납��A�`����[��Caltha dionaefolia�i�H
���A���̃n�G�g���\�E�Ɏ��������E�L���J�j�Ƃ����A�����m���Ă��āA�H���A���ł͂Ȃ����Ƃ͂��łɂ킩���Ă���̂ł����A���̊�Ȍ`���ǂ����Ăł���
�̂��s�v�c�Ɏv���Ă��܂����i�}�P�j�B���܂��܁A�`����[�̃i�o���m���ŃR�P�̍��ۊw��������̂ŁA���łɎ����n�����ɍs���܂����B���̃T���v�����
���ώ@����ƁA���Ԃ�A�t�̈ꕔ���Ȃ����Ė����������̂��ȂƎv���Ă��܂����B�܂��A�ߏ��ɕʂ̎�ނ�Caltha appendiculata��
������ނ������Ă��āi�}�Q�j�A������͂���ɖ�����������ŗt�̏ォ��t���������悤�ɂȂ��Ă���̂��悭�킩��܂����B���ꂼ��ȑO�ɕW�{�Ō�������
�͂������̂ł����A�����n�ł��낢��Ȕ����i�K���̕ψق̂���W�c������ƁA������W�{����������̂Ƃ͂��Ȃ������l�����U������Ă��܂��B��ɂ�
�����ӂ������ԎR�̏�ŁA�ӂ��Ǝv���o�����̂������t�c�o�L�ł����B���̉��|�i��A���i�͗t�悪���\���^�Ȃ̂ł����i�}�R�j�A�Ƃ��ǂ��A�t�̐�[�ɂ�
����t��t�����悤�ȗt���`�����܂��i�}�S�j�B����A�ǂ����A�ꂽ�t�̉����������Ƃ��̂悤�ȕ\���^�ɂȂ��Ȃ����Ǝv���Ă����̂ł����A����
�E�L���J�̗t�̖����ƈꏏ�ɍl����ƁA�A���̐V�K�`���i���ɂ���������̖����ׂĂ݂����Ȃ��A�Ǝv���Ă��܂����B����ɁA�H���A���̃E�c�{�J�Y���̓�
�R�ψّ̂ő܂������ɊJ���Ă��܂����̂��������Ƃ�����A���������Ă���������Ői�������̂��ȂǂȂǁA���낢��Ȃ��Ƃ��v���o����Ă��āA�����킸�A
�ɂ�ɂ₵�Ă��܂��܂����B����ɁA�����̍זE�ړ����āA�v�́A�זE���m�̐ڐG��ς��邱�Ƃ����ǁA�A���̖������č��܂ŐڐG���Ȃ������זE���ڐG���ĐV
�����`���ł��邱�Ƃ�����A���́A�d�g�݂Ƃ��Ă͓������Ƃ����A�ƋC�Â��A�����킸�A���������A�ƌ��ɏo���Ă��܂��܂����B
�@����A�A�����ٓI���Ȃ��Ďv���锭���l�������邩�ȂƎv���܂��B�i�o���m���ł͓d�q���C�����ǂ߂Ȃ������̂ł����A�g�����W�b�g�̎��ɁA���܂��܉^�ǂ�
�k�ĂɎ�������H���A���T���Z�j�A�i�}�T�j�̕ߒ��t�`���̘_���̃��r�W�������ʂ��͂��A��X�������Č������A�Ή���M�����҂̕�����������ɂ��肢���܂�
���B���̘_���iFukushima et al.
accepted�j�́A�T���Z�j�A�̕ߒ��t�͗t����̓���̏ꏊ�ōזE����������ω������邱�Ƃő܂��ł���Ƃ����A�זE�����ʐ��䂪�܂��܂���Ĉ�
�Ȃ����K�w�ł̑傫�Ȍ`�ԕω��������N�������Ƃ����������܂Ƃ߂����̂ł��B�A���זE�͓����Ȃ��̂ōזE�����ʂ̐��䂪��̌`�Ԍ`���ɑ傫�ȉe����^����
���B�����ł��זE�^��������Ă���g�D������̂ŁA���Ԃ�A�����悤�Ȏd�g�݂Ői�������튯�������Ƃ��肻���ł��B�����A���̂悤�ɕ���O�̂��̂��A
�����S�̂̕������������̂͂قƂ�Ǖs�\�ł��B����ȂƂ��́A���m��ɑ��k����̂���ԁB����DNA������́A�ӂ���͐ړ_�̂��܂薳�������W�̐搶
���ɂ���ł��A���낢�닳���Ă��炦��̂ŁA���������̂Ȃ��m�I�D��S�[���̋@��ɂȂ��Ă��܂��B���ЁA����Ƃ���낵�����肢�\���グ�܂��B
Fukushima, K., Fujita, H., Yamaguchi, T., Kawaguchi, M., Tsukaya, H.,
and Hasebe, M. Oriented cell division shapes carnivorous pitcher leaves
of Sarracenia purpurea. Nat. Com. Accepted.
Ohshima, I., Tanikawa-Dodo, Y., Saigusa, T., Nishiyama, T., Kitani, M.,
Hasebe, M., and Mohri, H. (2010). Phylogeny, biogeography, and
host-plant association in the subfamily Apaturinae (Insecta:
Lepidoptera: Nymphalidae) inferred from eight nuclear and seven
mitochondrial genes. Mol. Phylogenet. Evol. 57: 1026-1036.
Tanikawa-Dodo, Y., Saigusa, T., Chiba, H., Nishiyama, T., Hirowatari,
T., Ishii, M., Yagi, T., Hasebe, M., and Mohri, H. (2008). Molecular
phylogeny of Japanese skippers (Lepidoptera, Hesperiidae) based on
mitochondrial ND5 and CO1 gene sequences. Trans. Lepid. Soc. Japan 59:
29-41.

�}�P Caltha dionaefolia

�}�Q�@Caltha appendiculata�̂P���̗t�B

�}�R�@�����t�ւ̐�[���ꂱ�t

�}�S�@�����t�ւ̗t�̐�[�ɂ�����t���ł����悤�ȗt
�}�T�@�T���Z�j�ASarracenia purpurea�̂P���̗t
|
���q�A����Drymis�ɂ����x�I
�`���암�ɂ�
���J���@����
�@
��b�����w�������@�����i���������勳��
|
2014.07.08
No.15 �x�j�V�W�~�ނ̓�
���@����
�@�x
�j�V�W�~�Ƃ���������������
���낤���B������c���̘H�T�Ȃǂł悭�����鏬���Ȟ�F�̃V�W�~�`���E�ł���B���̒��Ԃ͐��E���炨�悻100�킪�m���Ă���A���̑����͑������ł�
�邪�A�M�ѓK���̎�ł͐X�ѐ��̑�������B�c���͎�Ƀ^�f�ȐA����H���Ƃ��邪�A�k���ł͑S���މ��̂Ȃ��A���Ɉˑ�������̂�����B���`�����͂��Ȃ葽
�l�ŁA�ꕔ�ł͋[�Ԃ������A���������̑��l���͗Y����퓙�̌`�Ԃ̕ω��Ƃ͂����Ί֘A�������Ȃ��B�܂��A�x�j�V�W�~�ނ̕��z�͂��Ȃ���قŁA���
���[���V�A�嗤��k�ĂɎY������̂́A�{���l�I�A�p�v�A�j���[�M�j�A�A���āA��A�t���J�암�A�j���[�W�[�����h�̂悤�Ȓn��ɏ����̎킪�u�����z���Ă�
��B�Ƃ��낪���̃O���[�v�̓�͂��ꂾ���ł͂Ȃ��B��L���������`�E����ƗY�����Ƃ̊W�����łȂ��A�����̌`�ԕω��ƒn���I���z�Ƃ̊֘A���������S
���Ȃ����̂���ŁA�n���I�Ȍq���肪�����ς茩���Ă��Ȃ��̂ł���B
�@���̕s�v�c���ɍŏ��ɋC�t�����̂́A�u�\����`�����w�v�Œ����Ȍ́E�ĒJ�čO�搶�ł������B1974�N�ɐ搶�͂���܂Ńx�j�V�W�~�ނ̕��z���S���m���
�Ă��Ȃ������p�v�A�j���[�M�j�A����V��Melanolycaena��
�n�݂���2�V��M. altimontana��M. thecloides��
�L�ڂ��A���킹�Đ��E�̎�ȃx�j�V�W�~�ނɊւ��鍂�����ނ̍Č������s�����B���̐܂ɖ{�Q����`�Ԃ�ԁA�n���I���z�Ƃ̊֘A�ł��̂悤�Ȕ�߂������
���o���A�𖾂ł���u�i���v��u�K���v�A�u�[�ԁv�Ƃ�������ʓI���������̂ɎQ�l�ɂȂ�ƍl�����i�ĒJ,
1999�j�B���̖������炯�̃O���[�v�ɂ����Ƌ��������������Ă���ꂽ�悤�����A���̓�������������Ȃ��܂܂ł������B
�@1998�N�ɏC�m�@���Ƃ��ċ�B��w�̑�w�@�i�w�������́A�����̐��b�l�w�������ł����������̌`�Ԋw����ьn���w�̌��ЁA�O�}�L���搶����̊���
�ŁA���̃x�j�V�W�~�ނ̍������ނ̍Č����ƌn���i���w�I�Ȍ������C�m�_���̃e�[�}��I�B�������A�O�}�搶���ĒJ�搶���炱�̕s���ȓ���Ă���
�̂����������ł���B���̌������s���ɂ�����A�܂��͎ĒJ�搶����̏��������炤���ƁA���킹�Ė{�����ւ̋��͂�v�����邱�Ƃ��O�}�搶���炲�w������
���B�����A�ĒJ�搶�Ɏ莆�������Ă����̂��肢��Ԃ����莆�𑗕t�����Ƃ���A�Ԃ��Ȃ����J�Ȃ��Ԏ����͂����B�����ɂ͎�����̂��肢�Ɋւ�������ƁA
�����g�̃R���N�V��������������Ă��镺�Ɍ����l�Ǝ��R�̔����قɑ����^��ŕW�{�������������Ƃ�������Ă������B���̌�A����ɓ��̎莆�Ɠd�b�̂�
����𑱂��A�ĒJ�搶�ƕ��Ɍ����Œ��ڂ�����Ė{�����̖��ƓW�]���������������B�ĒJ�搶�ƌ����A�x�j�V�W�~�ނ����邱�ƂȂ���[�t�B���X�̌���
�ł���������z���A�t�W�~�h���V�W�~�̑����ɂ���������Ă���`���E�ތ����̑�Ƃł���B��������ۏo���̃`���E�����������ɂƂ��āA�ĒJ�搶�͍\����`
�����w�����������_�w�҂Ƃ��Ă����A�ނ���`���E�ނ̕��ފw�҂Ƃ��Ă�������ْ��Ɗ������̕��������������Ƃ������Ă���B
�@��N����A���͏C�m�_�����Ȃ�Ƃ��܂Ƃߏグ�A�x�j�V�W�~�ނ̌n�����ފw�I�����ɂ��Ă͈ꉞ�̋�肪�������A���͌n���̕����ł͂������肱�Ȃ���
�a�����������B�܂�A�`�ԃf�[�^��p�����n����̓\�t�g�ɂ��Őߖ�@�œ����ꂽ���ʂɂ��܂莩�M���Ȃ������̂ł���B����ł��A�ĒJ�搶�ɂ͂�������
�Ă��ӌ����f�������Ǝv���A���҂ƕs�������荬�������C�����ŏC�_��o�̑O�ɂ��̓��e��X�������B�Ԃ��Ȃ����āA�ĒJ�搶�����ʂ̎莆���͂����B������
�́u�E�E�E���̂����p�l�ɂȂ�̂ł͂Ȃ����ƁA�����ꂨ�̂̂��Ă��܂��B�E�E�E���������\�͂�����������܂��B�������݂܂���A����ȏ�̓����̂悤
�ł��B�v�ƋL����Ă������B����܂Ŏ莆�̂�����搶�Ƒ����Ă������ŁA�������ɂ��̂悤�ȕ��͂͌������Ƃ��Ȃ��A�����ɐ搶�̑̒���S�z����C����
�����ݏグ���B���̍��ɂ́A���łɋC�͂�̗͂������Ă����̂ł��낤�B���̈���ŁA�搶�̂����҂ɂ������o���Ȃ������̂�������Ȃ��Ƃ������]�����P��
���B�搶�̂��l���ȏ�̂��̂�E���Ă��Ȃ������̂ł��낤���B
�@���̌�A���m�ے��ɐi���́A�e�[�}���V�W�~�`���E�ȑS�̗̂c���`�Ԃƌn���w�Ɋւ��錤���ɍL�������߁A�x�j�V�W�~�ނ̌���������ɒNj����邱�Ƃ͂�
�������B����ł��n���W�̈�a����s�����͂����Ɛ@���Ȃ��������߁A�@�����ΐ���Ƃ����q�n���w�I�ȃA�v���[�`����`�Ԃœ���ꂽ���ʂ���������
�ƍl���Ă����B�w�ʎ擾��A���͊�b�����w�������̒��J�����א搶�̏Љ�ŁA���Y�L�ނ̕��q�n���w�I�����Œm���铌����w�̏㓇��搶�̂Ƃ���Ƀ|�X�h
�N�Ōٗp���ꂽ�B�Ζ����ԊO�͎��g�̌��������Ă��ǂ������̂ŁA����̓`�����X�Ƒ����x�j�V�W�~�ނ̕��q�n����͂��n�߂邱�Ƃɂ����B���̎��̂��߂�
DNA�T���v�������͗��ߑ����Ă����̂ł���B�~�g�R���h���A�Ɗj�̑o����DNA����n����͂�i�߂Ȃ���A�s�����Ă���T���v�������߂ăA�t���J�⒆��
�O�@�e�}���Ȃǂɂ��K��A�悤�₭�x�j�V�W�~�ނ̍����n���ɂ�����S�̑����������̂́A�{�����̒��肩���10�N��̂��Ƃł������B
�@���ʂ��o�Ă݂�A�n���n���w�I�Ȋϓ_����ł����[�Y�i�u���ɉ��߂ł�����̂ł������B�Ⴆ�A�p�v�A�j���[�M�j�A�ɐ�������Melanolycaena�������̔����O�r跗�߂̌`�Ԃ��璆�Ă�Iophanus���ɋ߉���������Ȃ��Ƃ���Ă������A���q�n���ł͂ނ���O
�ς��قȂ�j���[�W�[�����h�̃O���[�v�Ǝo���Q���`�����A����ɑo�������킹����Q�̓`�x�b�g�⒆�������̍��n�ɐ�������Helleia��
�ɋ߉��Ƃ����i�W���iprogression
rule�j�łقڐ����ł���B�ł́A���q�n����͂ɂ�錋�ʂ����ۂɐ��������ǂ��������ƂȂ邪�A������Č`�Ԃ��������ƁA�s�v�c�Ȃ��Ƃɍ��܂ŋC
�t���Ȃ��������L�h���`�������X�ƌ����n�߂��B���̂悤�ɕ��q�n���̐�������������Ɠ����ɁA�`�Ԃ̎��ʂ╽�s�i���͑z�����ĒZ���ԂŋN���邱�Ƃ���
����A��d�̋����������オ�����̂ł���@�B
�@�ĒJ�搶�̐��O�ɂ��̌��ʂ������������������̂����A���ꂪ����Ȃ��̂��B��̐S�c��ƂȂ����B�܂��C�̂��̂Ƃ��R�̂��̂Ƃ��o���Ȃ����̏C�m�@��
���������ɁA�����̂��Ƃ��������������������łȂ��A����܂ŏ������߂Ă����V�W�~�`���E�Ȃ̗Y�����̌��}�܂ł�搶�͎��ɑ����ꂽ�B�ŋ߁A�����̕`
�}�߂ẮA�{����鎑�i�����邾���̌����҂ɐ���Ă��邩�ǂ������������g�ɖ₤�ƁA�܂������������Ă��Ȃ����ƂɋC�t���A�Ȃ���w�̓w�͂��K�v
�ł��邱�Ƃ�F������̂ł���B
|
���@����
�@
������w�������������ف@����
|
2014.02.07
No.14 �@���A�ɓ��������ʔ���
�r�J�@�M�Y
�@
�������߂ĕ��ƈɓ������̔��䓇�ɍ̏W�ɖK�ꂽ�̂́A������30�N�ȏ�O�̒��w�R�N�̉ċx�݂������B�����A�J�~�L�����V�ɑ傫�ȋ���������Ă������ɂƂ�
�āA�n�`�W���E�g�Q�E�X�o�J�~�L����n�`�W���E�R�o�l�J�~���ȂǑ����̔��䓇�ŗL�̃J�~�L�����V�̒��i�Ƃ̏o��͂܂��ɋ���̘A���������B�������A����
�����Ă��̍K���ȏ͈�ς���B�@
�u���̓��ɂ̓m�R�M���N���K�^����R�����B�v�h�̂���l���炻�̈ꌾ�����̂͏����̗[�H�̎��������B���̎��ɂ́A�{�y���炱��Ȃɗ���
�����ɂ��m�R�M���N���K�^������̂��Ə��X�����͂������A�V�����{�Ƃ����N���K�^���V�̏W�̃x�X�g�V�[�Y���Ǝv��ꂽ�����ł����������ɁA��R����̂ł�
��Α؍ݒ��ɊȒP�ɍ̏W�ł�����̂ƁA�����A���������Ă����B�������Ȃ���A��̓��Ό����ł��m�R�M���N���K�^�͑S���p�������Ȃ��B�R���ڂ̒��Ԃɓ���
�ōł��ǍD�ȐX�ъ����c���Ă���O���R�̒������т��ѓ���ŁA�m�R�M���N���K�^�Ǝv�������̈ꕔ���E�����B�����Ȃ�Ɛ�����ł��������m�R�M���N���K
�^�̎p���������Ȃ����B�Ō�̔ӂ͖ڕW���m�R�M���N���K�^��{�ɍi���ĕK���̓��ΒT�������s�����i���͒����ł��Ȃ��̂ɁA���̂��߂ɖ�ʂ��Ń����^�J�[��
�^�]����H�ڂɂȂ����j�ɂ��ւ�炸�A���ʂ͎S�s�ɏI�����B�@���������̒��A�ǂ����Ă����߂���Ȃ����́u�Ō�̓q���v�Ə̂��āA�a�镃��������A�h����
�킴�킴���������Č��̎O���R�̗ѓ����ċ�`�Ɍ������Ă�������B�������A�肢���ނȂ����O���Ƀm�R�M���N���K�^�����̈ꕔ���E�����ꏊ���߂��Ă�
�����ڗ��������ʂ͂Ȃ��B��������A��`�̂����ꋽ�̏W����������ƕ��͎Ԃ̃X�s�[�h���グ���B�ƁA���̎��A�����^�J�[�̎ԑ�����H��ɂЂ�����Ԃ���
�����N���K�^���V�̎p���͂�����ƌ������B�u�����I�v�������Ԃ��������A���͎Ԃ���~������������h�A���J���ĊO�ɔ�яo�����B�Ō�̍Ō�ɂ���Ƃ̎v
���Ŏ��ɂ������m�R�M���N���K�^�̏��^�̗Y���E�����Ƃ��ł����u�Ԃ������B���̎��̋����͍��ł��͂�����Ɗo���Ă���B�������P���̂��������ɂ��������^
�̗Y�ł͂��������A���܂�ď��߂Č������䓇�̃m�R�M���N���K�^�͐^�����ŁA�܂��Ɉٍ�������Ղ�A����e����ł����{�y�̃m�R�M���N���K�^�Ƃ͑S����
�Ȃ������݂Ɍ������B
�@���̌�A�w�Z�̐}���قɂ������k���فi���a38�N���s�j�́u���F���{�����嚤�ّ�Q���i�b���ҁj�v�ׂĂ݂�ƁA���̔��䓇�̃m�R�M���N
���K�^�͍̏W���̒����ǂ���A�n�`�W���E�m�R�M���N���K�^ssp.
hachijioensis�i�����̎��ɂ͂��̕\�L�̈Ӗ����番����Ȃ������j�ƌĂ�锪�䓇�ƎO��ɂ����Ȃ��ɂ߂ē��ʂȂ��̂ł��邱�ƁA���̈��
�Ŗ{�y�̃m�R�M���Ƃ̐��Ԃ̈Ⴂ�ȂǂɊւ��Ă͑S���𖾂���Ă��Ȃ����Ƃ�m�����B�u���������̓��ʂȃm�R�M���N���K�^���v�������ɍ̏W���āA���̐��Ԃ�
�𖾂������I�v����ȗ��A���̔��䓇�w���n�܂����B
�@�K�����̎��Ɓi���m���j�̗��e�͕��C��`�Łi����������ƕ��͐�̔��䓇�ł̌o���Œ��肽�̂�������Ȃ����j�A�����N�̎����P�ƂŁA������
�w�����Ɂi�傫�Ȑ��ł͌����Ȃ������R�w�Z���x��Łj�����̍̏W�n�ɒʂ����Ƃ��ٔF���Ă��ꂽ�B���A�Œ��w�E���Z����̖K����ʂ��āA�����̃n�`�W���E�m
�R�M���N���K�^���̏W�ł��������łȂ��A�����̔��������͖{�y�Y���������Ƒ����T�`�U���ł���{�y�ł̌o������x�X�g�V�[�Y���Ǝv���Ă����V�����{�ł�
�x�߂������ƁA���s�X����������Ԃɂ��������邪���ɂ͏W�܂�Ȃ����ƁA�c���͌͂ꊔ�̍�����|�̐ڒn�ʂɐ��E���邱�ƁA�Y���|�̉��œ꒣���
�Y���ɂ��鎓�ƌ�����邽�߂ɑ҂��\���Ă��邱�ƁA�Ȃǃn�`�W���E�m�R�M���N���K�^�Ɋւ��鐔�����̐V���Ȓm���邱�Ƃ��ł����B�n�`�W���E�m�R�M���N
���K�^�ȊO�ɂ��A�n�`�W���E�l�u�g�N���K�^�ⓖ���܂��L�^�̂قƂ�ǂȂ������q���^�N���K�^��R�N���K�^���̏W���A���Љ�i�Ƒ������j�𑗂�`�r�N���K
�^�̋����[�����Ԃ��𖾂��邱�Ƃ��ł����B��w���ɂȂ��Ă���͔��䓇�����łȂ��A�䑠����O��ɂ��n��A�~�N���~���}�N���K�^��}���N���K�^�̍̏W��
���������B
�@���Ȃ݂ɁA���ɂƂ��Ă̏��߂Ă̕ł���u���䓇�ɂ�����L�{�V�J�~�L���̍̏W��v�Ƃ����Z��i�����ނ� No.
190:21-22.�j�́A���̍��ɔ��䓇�œ���ꂽ�W�{�Ɋ�Â����̂ł��������A�n�`�W���E�m�R�M���N���K�^��`�r�N���K�^�̐��Ԋώ@�̌��ʂ́A���̌�
�̏C�m�_���u�N���K�^���V�ȍb���̔ɐB���ԁv�̎�v�����ƂȂ����B�������Č���ƒ��w�`��w���{�ے��܂ł̍ł������Ȏ����Ɍo���������̈�A�̈ɓ������w
�́A�܂��ɂ��̌�̍��������҂Ƃ��Ă̎��̌��_�ƂȂ����ƌ����Ă��ߌ��ł͂Ȃ��B
�@80�N��㔼����90�N�㔼�ɂ����āA������N���K�^���V�u�[����w�i�ɁA�ɓ������̃N���K�^���V�Ɋւ��镪�ފw�I�Ȍ������}���ɐi�݁A�n�`�W���E
�q���^�N���K�^��n�`�W���E�R�N���K�^�A�C�Y�~���}�N���K�^�Ȃǂ̐V�^�N�T�����X�ɋL�ڂ��ꂽ�B�n�`�W���E�m�R�M���N���K�^�Ɋւ��Ă��A���̓Ǝ��̌`�Ԃ�
���ԓI��������u�䑠���Ȗk�̖{�y�n�m�R�M���N���K�^�Ƃ͕ʌn���̔��䓇�ŗL�̓Ɨ���v�Ƃ��鋻���[�������������ꂽ�B����A���̍��̎��͐�痮����
�}���[�V�A���͂��߂Ƃ���C�O�̒����ɖv�����A�ɓ������Ƃ͂�������a���ɂȂ��Ă����B
�@�������A���N�O�A�v��ʓW�J���玄�̒��̈ɓ������ւ̏�M���ĔR���邱�ƂƂȂ����B���������̓~�g�R���h���ADNA��p���������̃m�R
�M���N���K�^�ނɊւ���n���n���w�I��͂������B��͂ɍۂ��āA���B�̈�����܂ޖ{�y�e�n�̃m�R�M���N���K�^�ƃn�`�W���E�m�R�M���N���K�^�����������A
���ʂ�O�A���́A�n�`�W���E�m�R�M���N���K�^���{�y�̃m�R�M���N���K�^�Ƃ͕ʌn���̎�ł���A����Έ⑶�ŗL��Ƃ��Ĕ��䓇�Ɋu�����z�������̂ł���
���Ƃ����q�ł����t������Ɨ\�z���Ă����B�n�`�W���E�m�R�M���N���K�^�̎o���Q���Ƃ��āA���̌`�Ԃ̗ގ������璆�����p�ɕ��z����}���o�l�m�R�M��
�N���K�^��z�肵�Ă������ł���B���������ʂ͗\�z�O�̂��̂������B�n�`�W���E�m�R�M���N���K�^�͈ɓ������̈ꕔ�̃m�R�M���N���K�^�̌Q�ƒP�n���Q����
���A���������̕���̒��x�͂��̑��̖{�y�Y�m�R�M���N���K�^�̒n��̌Q�Ԃ̕ψقƑ卷�Ȃ��������̂������̂��B�����Ď��{�����n�`�W���E�m�R�M���N
���K�^�Ɩ{�y�Y�m�R�M���N���K�^�̏ڍׂȌ`�ԑ���w�I��͂̌��ʂ�����A�n�`�W���E�m�R�M���N���K�^�ɂ݂���`�Ԃ�Ԗʂł̓��ꉻ�́A���t�Ƃ������
�I�ȉa���ԂŌ��܂�����K�v���Ȃ��Ȃ������߂ɁA�R�X�g�̂������A�S�̐L���������߂Ă��܂������ʂƂ��ċ}���ɐ��������̂ł��낤���Ƃ���������
���i�����̉�͌��ʂ̏ڍׂ�2009�N���s�̍���DNA�j���[�X���^�[NO. 2�ŏЉ�Ă���j�B
�@�܂����A����܂ł����͂��Ȍ̂����̏W����Ă��炸�A�قƂ�ǒm���̓����Ă��Ȃ������O���䑠���̃m�R�M���N���K�^�i�����̏W����
���炸�A���R�A��L�̕��q�n����͂ɂ��g�p�ł��Ă��Ȃ��j����������A���ꂼ�ꂪ�{�y�̌̌Q�Ƃ͕ʈ���Ƃ��ċL�ڂ����Ƃ��������i�H�j�����������B��
���Ȃ�ƎO���䑠���̌̌Q���܂ވɓ������S��̃m�R�M���N���K�^�̕��q�n���n���w�I��͂����{�����ɂ͂����Ȃ����A�~���}�N���K�^��q���^�N���K
�^�A�R�N���K�^�Ȃǂ̉�͌��ʂ��傢�ɋC�ɂȂ��Ă����B�K���A�������o�傹�˂Ȃ�Ȃ������̂Ƃ͈���āA���̓w���R�v�^�[�H������������ĒZ������ň�
�������e���ɏo��������B���ꂩ��͌����̍��Ԃ�D���āA�������ɂ͌��n�؍ݐ����ԂƂ������d�����ŁA�e���̒����Ɖ�̓T���v���̍̏W���J��Ԃ����Ƃ�
�Ȃ����B
�@�������ē���ꂽ�T���v���Ɋ�Â��ɓ������̃N���K�^���V�Ɋւ��镪�q�n���w�I��͂́A���݁A�܂��i�s���̒i�K�ɂ��邪�A�\��I�ȉ�͂���
�͗\�z�O�̋����[�����ʂ������������Ă���B���ɁA�䑠���Ɛ_�Ó��ɂ����������Ă��炸���E�I�Ɍ��Ă����M�����ɓ������̌ŗL��Ƃ��ėL���ȃ~�N���~
���}�N���K�^�̉�͌��ʂɂ̓n�`�W���E�m�R�M�����z����������������B
�@�~�N���~���}�N���K�^�͏��^�̃~���}�N���K�^�œ����̒��ł͑c��I�Ȏ�Ƃ݂Ȃ���Ă���B�`�ԓI�ȓ������璆���암�̉��͗���ɕ��z���郉
�G�g�D�X�~���}�N���K�^��p���[�~���}�N���K�^�Ɠ�����Q�Ɋ܂܂��Ƃ���A�]���̍l�@�ł́A�~�N���~���}�N���K�^�́A�c��킪�����嗤������{�A������
�ɓ������ւƐN��������A���{�{�y�ł͉����̗��R�Ŗł�ł��܂��A�ɓ������̌䑠���Ɛ_�Ó��݂̂ɐ����c�����A������T�^�I�Ȉ⑶�ŗL��ƌ��Ȃ���Ă�
���B���ۂ�DNA��͂̌��ʂ�����{�킪����璆���Y�̎�ɋ߉��ł��邱�Ƃ����t����ꂽ���A���̈���ŁA���ɋ����[�����Ƃɒ����Y�̋߉���ƃ~�N���~
���}�N���K�^�̈�`�I�ȋ����͋����قNj߂��A�~�N���~���}�N���K�^�̑c���̈ɓ������ւ̐N���i���z�g��j�͂����ŋ߂ɐ������\�����������ꂽ�B��X��
���̌��ʂ���A�~�N���~���}�N���K�^�̑c���͈ɓ������Ɋu�����ꂽ�̂ł͂Ȃ��A�É��͂̑�×��ɂ���Ĕ�r�I�V��������ɒ����嗤����ɓ������ɕY����
���\�����������ɂ�ŁA���݁A���̗��t���̂��߂̌������݂Ă���B
�@�����āA�ɓ������ɂ́A�n���n���݂̂Ȃ炸�A�i�������w�I�ɂ����ɋ����[����ނ��L�x�ɂ��邱�Ƃ����炩�ɂȂ��Ă����B�Ⴆ�A�����̉�
���ňɓ������̃N���K�^���V�ł͌ŗL�^�N�T�₻�̋߉����ތQ�̍\�����D��킪�����ɈقȂ�A���������ꂼ��̌ŗL�^�N�T�̐��Ԃ���j�����Ԃő傫����
�Ȃ��Ă���Ȃǔ��ɋ����[�����ۂ��ώ@����邱�Ƃ����������B�����������Ԃ̌Q�W�\���̍��ق�e���̌̌Q�ɌŗL�ȓK���`���̐i���ɂ́A���R�I���͂���
���A�Y�����͂��ߋ����I�ȕ��U�ɂ���ĐN�������c��̌Q�ɓ������n�n�Ҍ��ʂ��`�I�����A����ɂ͈�`�q�������Ԃ̋����ȂǗl�X�ȗv�������G�ɗ���
�����Ă���炵�����Ƃ����肳�ꂽ�B�܂��A�Â�����c�_�̂������Èɓ��������ɑ����āA�ŋ߂ł̓t�B���s���C�v���[�g�̖k��ɔ����ɓ������̊�̖̂{�B
�ւ̏Փ˂ƒO��R�n�̌`���ȂLjɓ������̐����n���w�ɖ��ڂɊW����n���Ȋw�I�Ȓm�����~�ς���Ă������Ƃ�����̈ɓ������̐������Ɋւ��錤����傫��
�i�W������v���ƂȂ邾�낤�B
�@���m��Ƌ��k��̋��E�Ɉʒu���A���m�̃K���p�S�X�ƌĂ��قǓƎ��̐����������ł���������A�ŗL��`�����̕�ɂƂ��Ė��������}
�������̏ꍇ�قǂ̔h�肳���Ȃ����߂��A�����Ɏ����ŗL�^�N�T�̕�ɂł���ɂ�������炸�A�ɓ������̍������Ɋւ���n���E�����n���w�I�Ȍ����͑傫
�������x��Ă��銴���ۂ߂Ȃ��B����ŁA�嗤�����f����Č`�����ꂽ�嗤���ł��问���̗l�X�Ȑ����Ɋւ���ߔN�̕��q�n���n���w�I��͂ɂ���āA����
����̓��̐������̌`���ߒ��͓��̐����̎����⏇�ԂƋɂ߂Ă悭��v���邱�Ƃ����炩�Ƃ���Ă���B�������A��������������������Ȃ����A����͂���Ӗ�
�\�z���꓾�铖�R�̌��ʂł���A���f�ƕ��U�i�����I�Ȋg�U�j�̑o�������ݍ����ɓ������̐������̐����̉𖾂͈�ؓ�ł͂����Ȃ��䂦�ɗy���ɖʔ����B����
�����嗤�����f����Č`�����ꂽ�嗤���ł��问�����ΎR���̊C�m���ł���K���p�S�X�ɂȂ��炦�鎖���̂��i���Z���X�ł���A���̓_�ł͂ނ���ɓ�����
�̕�����قǁu�i���̎�����v���铌�m�̃K���p�S�X�ƌĂ��ɂӂ��킵���悤�Ɏv���B
�@���̂悤�ɐ����w�I�ɔ��ɋ����[���ɓ����������A�ɂ߂ėR�X�������ƂɁA�ߔN�A�J���ɂ�鐶�����̔j��ɉ����đ����̊O���킪�N�����A
���̋M�d�ȍݗ����Ԍn�͉�ł̊�@�ɕm���Ă���B���s�X���̋����ɓ������̍����ނɂƂ��Ă̓C�^�`�ƃm�l�R�Ȃǂ̑�^�ٓ��ނɂ��ߐH�͉�œI�ȑŌ���^
������B�C�^�`��m�l�R�ȊO�ɂ��A�V�J��L�����A���M�̐H�Q�ɂ��X�ѐA���̔j����������B�O������Ɋւ��ẮA�����⏬�}���ł͂��̑�X�I
�ɍu�����Ă��邪�A�ɓ���������x��ɂȂ�O�ɁA�ꍏ���������炩�̑���Ƃ�K�v������B���̂��߂ɂ��A�{�e�����������ƂȂ��āA�M�d�Ȉɓ������̐�
�����Ɉ�l�ł������̕��������������A���̑��l���̉𖾂�ۑS�ɌW���Ă��������悤�ɂȂ邱�Ƃ�Ɋ肤����ł���B
|
�r�J�@�M�Y
�@
��B��w��w�@�@��r�Љ�������@�@����
|
2013.06.27
No.13 �@�`���E���̎���
�ї��@�G�Y
�@�O�ɂ����������Ƃ����邪�A���͎q���̍������
�u�`���E���v��
�͂Ȃ��B���ł�
�����ł͂Ȃ��Ǝv���Ă���B�������ɏ����������琶�����D���ł���A�ĂƂ��Ȃ�ΐ^�����ɂȂ��ăZ�~��g���{��L���M���X��ǂ������Ă������A�`���E��
�̂����莔�����肵���L���͂Ȃ��B�t�@�[�u�����Ă��Ă��A�����Ă݂����Ǝv�����̂̓Z�~��A���ł������B
�@���܂ꂽ���N�ɖ��B���ς��N����A����ł��R�����N�Ƃ��Ĉ������ɁA���e���E�ƌR�l�ł������̂ŁA���j�ł��鎄�͉��̋^�����Ȃ����̌�������߂ɗ�
�R�c�N�w�Z�ɓ������B���R���Z��{�����邽�߂̊w�Z�ŁA���w��N����N���I���ē��Z���A�O�N��ɗ\�Ȏm���w�Z�A����Ɏm���w�Z�i���̖h�q��w�Z�ɑ����j��
�i�ނ̂ł���B���������Z��N�ڂɔs��ƂȂ�A������̖ړI�͍�����v���K�^�ɂ��ʂ������ɏI������B
�@���͈�]���čD���Ȃ��Ƃ���낤�Ƃ������ƂɂȂ�A�������Z�ő�w�̗��w���ɓ����w�ȂƂ������̂����邱�Ƃ��͂��߂Ēm���āA����̓��w�Ȃɐi�w��
���B���Ƃ��čŏ��ɏA�E�����̂��O��̗ՊC�������ŁA�����ŃE�j�̐��q�Ŏ������n�߁A�Ȍ㐸�q���߂��鏔���Ɏ��g�ނ��ƂɂȂ����B���̊Ԑ��q�̕ږт�
��v�ȍ\���v�f�ł�������ǂ̎听����V�����^���p�N���Ɠ��肵�ă`���[�u�����Ɩ�������K�^�ɂ��b�܂�A���q�w�Ƃ����w�╪�����邱�Ƃ��ł����B����
�ƍ����͑��̐l�����̌����e�[�}���Ǝv���Ă����B
�@���ă`���E�Ƃ̏o��ł��邪�A���傤�Ǒ�w�����̂���ɉƓ��̐e�����쌧�̐������ɎR�����������B�s���Ă݂�Ƃ��ĕ��R�C���Y���́u�����}�Ӂv��
�����������Ƃ̂Ȃ������`���E��g���{�����������B�����Ŏ���̂��̂����ł��S���̂��Ă݂������̂ƁA�l�\�̎�K���ō����̏W������悤�ɂȂ����B�c
�O�Ȃ���̂�Ȃ��������A�����ɂ̓I�I�E���M���q���E�������܂������悤�ł���B���̌�I�[�X�g�����A�ɗ��w�����ۂɁA�����V�h�j�[�ɂ���ꂽ�ĒJ�čO��
��ɂ��낢��Ǝ�قǂ����A�`���E�ɂ̓|�C���g�Ƃ������̂����邱�Ƃ��w�B���̉��Ŕ��{�b�s����ⓡ���m�v����Ƃ��m�荇���悤�ɂȂ����B
�@������N�ő��Ƃ��ĕ�����w�ɍs������A�����ɂ̓`���E�̗ؕ��𑖍��d���Ŋώ@���Ă����u�`���E���v�̐V��@�ׂ������B�����Ń`���E�̐��q�̓d��
����肩�������A�Ԃ��Ȃ���b�����w�������̏����Ƃ��ĉ���ɕ��C���邱�ƂɂȂ�B��C�ɂ͑�w�̓������Ń~�g�R���h���A�̌��Ђł��钆��@���N�������B
���傤�Ǒ��V�ȎO�������������قŃ~�g�R���h���ADNA�ɂ��I�T���V�̕��q�n�����n�߂�����ł���B
�@�V�삳��͒��V�N�̒�q�̖q�c�T���N�Ƒ��V����Ƀe�N�j�b�N��������āA�M�t�`���E�̋N���ɂ��ă~�g�R���h���ADNA�ň�̓������o�����ƂɂȂ�B
���͊���̏����Ƃ��Ă����ς猤�����̉^�c�ɓw�͂��Ă������A�A���̕��q�n���̃p�C�I�j�A�ł��钷�J�����N������ɕ��C���Ă����@����Ƃ炦�A���V��
�����{����A���؍F�i�����Ƃ������āA�����܂��܂������������������̃V�[�P���T�[���������A�`���E�̕��q�n���̑S���I�ȋ��������𗧂��グ�邱
�Ƃɂ����B
�@���Ă��ꂩ�炪�{��ł���B���̃v���W�F�N�g�ɂ͑S���́u�`���E���v���������͂��Ă��ꂽ�B�����\���N�O�̂��ƂɂȂ�̂ō����͂܂��܂�������
���A���łɑ����̃`���E�����̓V�R�L�O���ł�������A��Ŋ뜜��ł������肷��̂œ��{�Y�̂��̂��W�߂�̂����J�ł������B���Ɉꎞ�͂��ꂪ�Ō�̕W�{
�ɂȂ邩���m��Ȃ��Ǝv��ꂽ�I�K�T�����V�W�~�̍̏W�ɂ��Ă͊��ӂ�����Ȃ��B����ő����́u�`���E���v�̕��X�ƃR�l���������Ƃ��ł����B�������̎���
��������������Ȃ̍̏W�����Ƃ�̂ɑ傢�ɋ��͂��Ă���A�S���犴�ӂ��Ă���B
�@������N�őނ��ĊԂ��Ȃ��A�O�}�L������̎����Ŗ�㏟�炳���ƃS�}�V�W�~�̕��q�n������邱�ƂɂȂ����B����܂ł͂ނ��������̏W�́u�`���E
���v�̊F����ɂ��C�����Ă����̂ł��邪�A�����ŏ��߂Ď��炠�������̏W�ɕ������ƂɂȂ����B���̍ŏ��͗����͂��߂Ƃ���Ìy�����ł������B�����ŃS
�}�V�W�~���̏W�������Ă���k�C���̎R�{��������Əo��A�v���Ƃ͂���Ȃ��̂��Ƃ��̌����ȍ̏W�Ԃ�ɂقƂقƊ��S�����B�`���E�̍̏W�Ɋւ��ẮA����
�����Ă݂āA��コ���R�`�̉i���ÔV�����̗͗ʂɂ��傢�Ɋ����������B
�@�S�}�V�W�~�͖k�C����X����������_�X�Ƃ��Ă������������A�������W�߂�̂���ςł������B���̏\�N�̊Ԃɂ��̂�Ȃ��Ȃ����ꏊ������B�܊p����ɂ�
���̂Ɉ��m���A�É����ł͂��̃v���W�F�N�g���n�܂�O�ɐ�ł��Ă��܂����̂����Ƃ����ɂ����B�����W�{����ł͂܂Ƃ���DNA�����o�ł��Ȃ�����ł���B
����ł��ǂ����嗤�̎������܂߂����͌��ʂ��߂������ɘ_���ɂȂ�Ɗ��҂����悤�ɂȂ����B
�@����Ƃ͕ʂɁA�ߔN�͎������I�ɂ܂��o����Ă��Ȃ��`���E�����߂Ă����炱����ɏo�����Ă���B�����̃`���E�̂قƂ�ǂ��䂪���̐�Ŋ뜜���n���
���ł���B�����ɂ����Ĕ��������u�`���E���v�̊F����̎��͂̂قǂɂ͂܂��ƂɊ��Q���ւ����Ȃ��B�s���|�C���g�ŏꏊ�⎞���������Ă��������̂ł���
���A�����̏ꍇ�������Ă���B�����̂͂��̏ꏊ�ɂ͕K���ƌ����Ă悢�قǕʂ́u�`���E���v�����邱�Ƃł���B��������V�����̃N���}�c�}�L�`���E��
���߂Ď�����߂��̃|�C���g�ɍs�������A�ɓ����v����ɑ������������łȂ��A�����ɓ�A�O���ʂ��Ă���Ƃ��������▼�É��̐l�A�����Ƃ���ꏊ�ő҂��\��
�Ă����҂Ȃǂ������B
�@��コ��̗U���Œ��ފw��Ɋ���o���Ă݂Ă��A���ނ�ԁA���̑��`���E�̂������Ɋւ���u�`���E���v�����̒m���A�����ɂ́A���X�Ȃ�����n��
�͑����ł��ł��Ȃ��Ƃ��Â��v���m�炳��Ă��鍡�����̍��ł���B�g��@���A����s���A�ɓ����v�Ƃ������i�H�j�u�`���E���v�̂����ꂽ���q�����w�҂���
������̂��A�q���̍�����ώ@���ʂ����낢��ƕ��́E�l�@���邭�������Ă������߂ł��낤�B����ł��u����DNA������j���[�X���^�[�v�ł́A������
�u�`���E���v�����ɍv���ł����̂ł͂Ȃ����ƂЂ����Ɏv���Ă���B
|
�ї��@�G�Y
�@
������w����ъ�b�����w�������@���_����
|
2013.05.20
No.12 �@�����ɂȂ�Ȃ��������ƍ����c�m�`������
���@�T��
�@�u�����v�́A���ɂƂ��Ă�������ƃR���v���b�N
�X�̓����
�������t�N�U�c�Ȋ������N���閂�@�̌��t�ł������B�q���̍��͍�������D���ł��������}�ӂ�����ĕ����u�����͍����w�҂ɂȂ�v�ƌ����Ă܂����
�����B�������A���w���̍��ɕ�e�Ɍ���ꂽ�ЂƂ��Ɓ\�\
�u���܂̍����w�҂͎E���܂���錤���������ĂȂ������߂Ƃ��Ȃ����v
�@���̈ꌾ�ŁA���͍����D���������w�҂̖����������Ă��܂����̂ł���i���Ȃ݂ɁA�����ƌ�ɂȂ��Ă��炻�̘b���ɂ����Ƃ���u���炻��Ȃ��ƌ�������
����v�Ƃ܂������o���Ă��Ȃ��悤�������j�B
�@�u�����v�ɕ��A����`�����X�����̌�Ɉ�x�����������B�k��ɓ��w�������́A�q���̍��̖����v���o���Ē����i����������j�̌˂�@�����̂ł���B����
���A�������������A�����������Â��i���X�ɘA��čs����Ēj�S�l�ŃR�[�q�[�������邾���̒n���Ȗ����i�����A�����͎����܂߂ĂS�l�������Ȃ������j�B
�u����ł́A���̐t�͏I���ȁv
�@�����v�������͑��X�ɑޕ����A�ȍ~�͒��Ƃ͖����̑�w�������𑗂邱�ƂƂȂ����B�������A�������Ɖ����������Ƃ����āA�Ƃ��Ɋy�����t���オ�K��
���킯�ł��Ȃ��A���͂����u�����v�Ƃ������t�ɑ��Ă��悢��[������ƃR���v���b�N�X���点���̂ł������B
�@�C�m�ے��܂Ŏ��͗L�@���w���w�B���̌�A�����w�ɓ]��������̊�b�����w�������ɐi�w�������A���m�ے��őI�e�[�}�͋��ނ̐��B�����w�������B��
�����A�����Ƃ͑S���W���Ȃ��B������A�ї��G�Y�搶�≪�c�ߐl�搶��啽�m�v�搶�Ƃ����X�S�������������߂��ɂ������Ƃɂ͂܂������C�����Ȃ���
���B�����ł��܂��u�����v�ɂȂ�`�����X���Ă����̂ł���B
�@���̌�A���É���w�Ɉڂ��Ă���A���������̌�������邱�ƂɂȂ����B���������Ȃ牽�ł��悩�����̂����A�����ăz�^�������C�t���[�N�̂ЂƂɑI��
�̂́A���Ԃ��ɑ��钷�N�̓���ƃR���v���b�N�X�̂����ł��낤�B
�@�R���c�L���V�ɂ������ނ�����Ƃ������ƂŁA�Ȃ�̏Љ���Ȃ��啽�m�v�搶�Ɏ莆�������Ă݂��B����ƁA�R���c�L���V�ɂ��đS���m���̂Ȃ���������
��ϐe�ɂ��w������������ɁA���V�ȎO�搶���Љ�ĉ�����A���傤�ǔ�����������̍���DNA������ւ̎Q�������߂Ă����������B���������Q������
�݂���A
�u����[�H�v
���B�����w�̕��ł͂悭�����グ�Ă����ї��搶��x���搶�������Ă����̂Ńr�b�N�������B���ӂ���Ƃ��B����������̂��i�B��Ă����킯�ł͂Ȃ��Ǝv��
���j�B
�@����DNA������́A���̂悤�Ȓ����R���v���b�N�X��������P�Ȃ郀�V�D����g�������Ԃɓ���Ă��ꂽ�B���ꂪ�������Ė���Q�����Ă��邤���ɁA���̌�
������䂪�z�[���O���E���h�ł���ł����S�n�̗ǂ��ꏊ�ł���悤�Ɏv���Ă����B����Ȗ�ŁA2010�N���{��̎����ǂ����������Ē������Ƃ�
�Ȃ����B�ق�Ƃ��́u�����v�ɂȂ�Ȃ����������ɁA���߂āu�����v�̊F����̂���`�����ł��邱�Ƃ����͂ƂĂ��������B
�@�{������ɂ́A���D�������̂܂ܐ��ƂƂ��������w�ҁA��������D��������ǂ����E�ƂƂ��Ȃ������Ȋw�ҁA��������Ƃ��Ă���A�}�`���A���D�ƁA����
�Ď��̂悤�ɒ����ɂ͂Ȃ�Ȃ��������������������Ă���Ȋw�҂ȂǁA���낢��ȃ^�C�v�̉�����������Ă���B�������A�S���̎v���͂ЂƂu�������D���v��
�̂ł���B�����Ă��݂����m�����܂��Ȃ����킹�Ă���̂��uDNA�v�̃L�[���[�h�ł͂Ȃ����Ǝv���i���̃L�[���[�h���Ȃ���A���̏o�開�͂Ȃ���
���j�B���̂��C�ɓ���̑f���炵�����̉�A���߂Ď��������ǂ𑱂��Ă������͑��������������̂ł���B
|
���@�T��
�@
���É���w��w�@�����_�w�����ȁ@���q�@�\���f�����O��������@����
|
2013.04.22
No.11 �@
�@ �R���� ����@��
��
�݁A���͕��Ɍ��O�c�s�̎O�c�s�L�n�x�m���R�w�K�Z���^�[�Ɋw�K�w�����i�O�c�s�����E���G�����S���j�Ƃ��ċΖ����Ă���B�W�����쐬�A���R�w�K�Ɋւ�闈��
�҃T�[�r�X�A���畁�y�����Ȃǂ���ȋƖ��ł���B���Z���^�[�̎�ȗ��َґw�͏��w���ȉ��̎q�ǂ��B�ł��邽�߁A�ł��邾��������₷�������́g�ʔ����h��
�`���邽�߂ɓ����Ђ˂���X�𑗂��Ă���i���Ȃ݂Ɏ��ȏЉ�ʐ^�Ɏʂ��Ă���͉̂ߋ��̓W���ɗp�����e�C�I�E�[�~�̕W�{
�ŁA���̊J���͂����ӂ��\�[�X�̗e
��Ɠ������炢�̑傫���ł��邱�Ƃɒ��ځj�B
�@�������̍D���̎q�ǂ������̂܂ܑ�l�ɂȂ����悤�Ȏ��́A�ޗnj��g��S���g�쑺�Ƃ����I�ɔ����̎R���Ő��܂������B2013�N
���݁A�l�����悻2000�l�̉ߑa�̑����B���̎��Ƌ߂��ɒ����\�����������Ă���A�������̋g
��n��͒J���[���n�`���}�s�ŁA�R�̎Ζʂɏ����ȏW�����_�X�ƕ��z���Ă���B21���I�ɓ���\���N
���o�߂������A���̎��Ƃł͂��܂��ɐd�ŕ��C���Ă���B�܂��A�Ăɂ͓����͖Ԍ˂����������J���������ɂ���̂ŁA�Ƃ̒��̌��ԂƂ������ԁA�Ⴆ�^��
�X�̈�����╨����̈����ˁA�_�I�Ȃǂ͎��I�̒��ԁA���Ƀn�G�g���O������郂���L�W�K�o�`�ɐ�̂���D�̑������������悤�ɂȂ�B���ɂ��삪���w
�Z�w�萅�j��ɂȂ��Ă��邱�Ƃ�A���ނ�̍Œ��ɖڂ̑O���J���l�Y�~���j���ł������̂Œǂ������ă^���Ԃł����������ƁA�ȈՐ����̎搅���ɃI�I�T���V��
�E�I���l�܂��Ă������ƁA�c�����X���b�p�𗚂����Ƃ����璆�Ƀ������K�������Ă������Ɓc���X�R���Ȃ�ł̘͂b�ɂ͖����ɂ��Ƃ܂��Ȃ��B
�@����Ȋ��ň�������͂ǂ������킯���A�c�������璎������ɋ���������A�ʉ��E�ʊw�̓����ʼn�����߂܂��Ă͌��ւ���̃v���X�`�b�N�P�[�X�ɓ���
��
��
�炵�Ă����B�c�t���̍��ɂ͑������u�ނ��͂����v�ƌĂ�A�l���ň�x�ڂ́g���m��(�ނ�)�h���擾�����B
�@�g��ƌ����X�M�̐A�т���ŁA�A���͂��܂�ʔ��������̂����A�W���̎��ӂɂ͏��K�͂ȃN�k�M�т��c���Ă���B���̎��Ƃׂ̗ɂ��N�k�M�т�����A����
����
�͂悭�g�Q���W�i�N���K�^�̂��Ɓj�h���̂�ɍs���Ă����B�ߋE�n���ōL��������n�������Ǝv���邪�A�g��ł̓N���K�^�̑��̂��Q���W�A�R�N���K�^
�̓w�C�P�A�m�R�M���N���K�^�̓X�C�M���E�A�~���}�N���K�^�̓}�N���ƌď̂��Ă����B
�@���w�Z1�N���̉āA���͈�l�ł����̃N�k�M�тɃQ���W���̂�ɍs���Ă����B��l���q�œ��N��̎q��
���̐������Ȃ��������Ƃ�����A�����̗V�ё���͒��������B�����Ă������悤�ɁA�ڐ���t�����N�k�M���R���ĉ���Ă�����E��Ɍ��ɂ��������B�X�Y
���o�`�Ɏh���ꂽ�̂��B���͂��܂�̋��|�ƒɂ݂ŋ�����߂��Ȃ���ƂɋA�����B�����̓L���J���őΏ����邭�炢�����Ȃ��A���̏����ȉE��̓p���p���Ɏ��
�オ�����B���炭�͎w���Ȃ��炸�������ĂȂ������̂ŃX�v�[���ł��т�H�ׂĂ����̂������Ă���B���̌ジ���ƁA���Z�����炢�܂Ńn�`�ɑ��鋰�|�S��
�Ȃ��Ȃ������邱�Ƃ͂Ȃ������B
�@���w�Z3�N�����炢�������Ǝv���A�Ƃ̒��Ŏ��]�Ԃɏ����K�����Ă����B��ɏq�ׂ��悤�ɕ��n����
���}�ȍ����Ȃ̂ŁA���]�Ԃ̗��K����悭�炢�����ł���ꏊ�����������B���x�������Ȃ���A���K���J��Ԃ��Ă���ƁA�O����傫�ȃn�`���������Ɍ�
�����Ĕ��ŗ����B���́u�A�J���I�h�����I�����ȁI�v�Əł��Ď��]�Ԃ̌�����ς��ĕK���œ����悤�Ƃ�����c�Ȃ�ƁA���]�Ԃ������ł���ł͂Ȃ����B��
�̎��A�n�`�ɒǂ��������Ȃ�������A���܂��Ɏ��͎��]�Ԃɏ��Ă��Ȃ���������Ȃ��i����Ȃ��Ƃ͂Ȃ��Ǝv�����ǁj�B
�O
��̃����[�R���������M���ꂽ�ē����q����͂��߁A���̒m�荇���̍����w�ҁA�����w�҂͓s��܂�̕��������B�����āA���ł������C��d�ŕ����Ă���Ƃ�
�����Ƃ͏o����������Ȃ��B��������ƍ�������ɑ��鋻���ɑ��ւ�����̂��ǂ����A���ɂ͕�����Ȃ��B���������l���䂪�傫�����Ă���̂�����A
���̊����Łg�������̍D���h����������̂ł���A�s��炿�̍����w�҂���w�҂������͓̂�����O���B
�@�O��̃R�����ŐG����Ă����ē����q����̋^��Ɠ��l�A����������̖ʔ����ċ����[�����Ƃ��q�ׂ�̂͗e�Ղ��̂����A�ł͉��̍D���Ȃ̂��A�Ƃ���
��
���ɂ͂�����l���Ă������邱�Ƃ��ł��Ȃ��B����͖{���ɓ�ł���B���ꂪ������Ύ����g�ɂ��Ă����Ɨ����ł���̂�������Ȃ��B�����Ă����ƍ����
��
�̖ʔ������q�ǂ������ɓ`���邱�Ƃ��ł��邩������Ȃ�
|

����@��
�@
�O�c�s�L�n�x�m���R�w�K�Z���^�[�@�w�K�w����
|
2013.03.20
No.10 �@
�@ �����يw�|���Ƃ��Ă��������� �ā@���@���@�q
����DNA������炱�̃R�����Ɏ��M�˗������������āA�܂Ƃ܂���DNA�̌������ʂ������͓̂���̂ŁA���������邩�ǂ��������܂������A�R����
�Ƃ������ƂȂ̂�DNA����O�ꂽ���ł����������������邩�Ǝv���A�����g�̂��Ƃ��������Ă����������Ƃɂ������܂����B�q���̍�����ǂ̂悤�ɒ��Ɋ�
����Ă������A�Ƃ������ƂƁA�������ق̊w�|���Ƃ��ē������œ��������Ă��邱�Ƃ����`�����Ă݂����Ǝv���܂��BDNA�̘b�肩�痣��Ă��܂��܂����A��
�����������������B
�@�����قō����S���̏����w�|���Ƃ��ē����Ă���ƂȂ��������D���Ȃ̂ł����H�ƕ�����邱�Ƃ��悭����܂��B�����Ă�����͓�ł��A�Ƃ��������Ă���
���B���e���Z�����ɐ������̂ɋ�������������ł��Ȃ��A������͓̂����̕i��Ƃ����A���Ɏ���ɒ�������������ł�����܂���ł����B����ł��Ȃ�
�����S�����������ŃZ�~�̂��������A�T���V���ɃA�Q�n�`���E�̗c����������Ɛ������y���݂Ɍ�������肵�Ă��܂����B���ł���I�I�X�J�V�o���n
�`���ƐM���āA�Ƃ̃K���X�˂̌��Ԃ���Ԃ��o���Ďh����Ȃ��悤�ɍ̂낤�Ƃ�����A�J�Ɍ����z�킹�Đ������R�b�v�ɓ���Ă����Ɛ��ʂɗ����Y�ށA�Ə�
���Ă���{�����āA�����̑������Ō����z�킹�����̂̋z����������Ă��܂������ƂȂǂ��A���w���̎��̓��Ɉ�ۂɎc���Ă���v���o�ł��B���̌�A���Ɏ���
�܂Œ��𑱂��Ă���ꂽ�̂́A���Ԃ�Ƒ��̋��͂ƕ��C�����������炾�Ǝv���܂��B���̂��߂ɒ|�̖_�ŕߒ��Ԃ̌q���Ƃ�����Ă��ꂽ�c���A�����V���`���E��
�H���̏u�Ԃ��ꏏ�Ɍ��Ă��ꂽ��Ƒc��A���e�Ɨ��s�ɏo�����Ă���ԁA�����Ă����A���W�S�N�ɃG�T�i�A���j������Ă��ꂽ�c��A�Ƃ������͎҂����܂����B
����ɁA�U���K�j�A�������A�w�r�̔����k�Ȃǂ܂ŁA���ȊO�ɂ��l�X�ȕ����Ƃ̒��Ɏ�������Ŏ��Ȃ��Ă������ɂ�������炸�A������������邱�Ƃ̂Ȃ�����
�Ƒ��́g���C�h�̂��A�ō�������ƁA��l�ɂȂ��Ă���C�Â��܂����B
�@�����̏��q���w�Z�ɓ��w���������l�Œ��𑱂��Ă��܂����B���w��N�̉ċx�݂̎��A���s�Ђ���悵�������̏W�̗��ɐe�q�ŎQ�����A���߂Ē��̓W���Ƃ���
���̂��K�����̂����������ɁA���̍̏W�ɂ͂܂�܂����B�u�V���������̏W�v���o�C�u���Ƃ��Ĕ�������O��q��Ȃǂ֍s���A�̂������Ƃ̖����`���E���̂���
�W�����邱�Ƃɖ����ɂȂ�܂����B�I�I�����T�L�����߂ăl�b�g�C���������̎�̐k���銴�������ł��ǂ��o���Ă��܂��B�̎����ɂȂ�ƁA���ɏ�����[��
�l���邱�Ƃ������A�Ƃɂ��������w�������̂����w�ɍs�������Ǝv���A���̌�܂��܂����ɂ̂߂荞�݂܂����B30�ōK�^�ɂ������قɐE�āA���݂܂�
�����ƒ��Ɗւ�邱�ƂɂȂ�܂����B
�@�����قł͎q���̎��R�����̑��k�������Ύ܂��B��������͎d�����Z�����̂ł��傤���A����Ă���͕̂�e�Ǝq���̂��Ƃ������ł��B�����Ă��q����
�قƂ�nj����J������e���肪����ׂ�܂��B�����Ďq���ɗ��h�Ȏ��R�����������������ǂ�����悢���A�Ƃ����ꍇ�������ł��B���̑��k���e�́A���Ƃ�
�A�����w�Z�֍s���O�ɉƂ̎���Œ��̃Z���T�X���q���ɂ��������A���N������������T�Ԃō��܂����A�ǂ�����悢���A�Ƃ��A�R���e�i�ɐ������r�I
�g�[�v������Ăǂ�Ȑ������̂����邩�ώ@���Ă������A�������N����Ȃ��̂Ń��_�J���Ă��ĕ���������ǂ���ς葼�ɉ����N����Ȃ��A���R�ώ@�ɉ���
�R�c�͂���̂ł��傤���A�ȂǂȂǂł��B�ċx�݂��c�肠�ƂP�T�ԂƂ��������ɂ���ė��āA�h��̎��R�����ɉ������Ηǂ����낤���A�Ƃ������k�������
���B������Ђǂ��o�����́A����ċx�ݎq�ǂ����k��ł̎��ł��B��e���q���̏h��Ə̂��ĎB�e�������̖��O�������ė~�����ƁA�R���p�N�g�f�W�J���̉t��
��ʂŎʐ^�̒������\�������肳�����Ă������̎��ł��B�ꏏ�ɗ��Ă����q���͂����ɖO���Ă��܂��A������ŃT���_���R����n�߁A���x�ڂ��ɏR��グ���T
���_�����ׂœ���Ή����������ʂ̐l�̕W�{���̒��Ɍ����ɗ������̂ł��B�������W�{�͉��A�����ł͂��܂���Ȃ��ƂȂ�܂����B��e���ʐ^�̒���
���O��t���邱�Ƃɖ����ł�������q���������̂��ƂȂ��������ł��B����Ȏ��͉��̂��߂Ɏd�������Ă���̂��킩��Ȃ��Ȃ�܂��B�����͋ɒ[�ȗ�ł�
���A�����ق֍����̑��k�ɗ���ꂽ���ꂳ��ɂ́A���������y�������Ƃ����܂��傤�A�ƌ����āA�قƂ�nj����J���Ԃ̖��������q���ɉ����D���Ȃ́H�ƕ���
�ƁA�N���K�^���D���A�A�����ʔ������A�Ȃǂƌ��\�������Ԃ��Ă�����̂ł��B
�@���̉Ƒ��͑O�q�̂悤�Ɏ����g���C�h���Ă����̂ŁA���h�Ȍ�������点�������甎���ق֘A��čs���A�Ƃ����悤�Ȃ��Ƃ�����܂���ł������A����ł���
�s�Ђ̍����̏W�̊��ɎQ�������Ă���܂������A���̌�̉Ƒ����s�͂������̌v�悵���̏W���s�ƂȂ�܂����B�܂莄�́A���e�́g���C�h�̌��ʁA�ʔ���
�Ɗ��������ɂ��Ă����ƒm�肽���Ȃ�A���ʂ̏��̎q���s�����������������w�����̂����w�ւ̓��w�������ꂸ�ɍ�������킯�ł��B�e�͎q����������
���������Ƃd���Ď菕�������Ă�����̂��ǂ��Ǝv���Ă��܂��B��������Β��ɂ͍����ɋ��������������A�����DNA�����ɓ���𖾂������ƍl����
��҂�����Ă����̂ł͂Ȃ����A���̂��������ƂȂ�Ɩ��N�ώ@��⍩���W�{�̂���������Ȃǂ𑱂��Ă��܂��B�����Ŏ��ۂɎq��Ă��������Ƃ��Ȃ���
�ł��܂�����͂͂Ȃ��A�q��Ă͂���Ȃɂ��܂������Ȃ��A�Ƃ�������邩���m��܂��A���ꂩ������炭�͊w�|���Ƃ��Ă����������𑱂��Ă���
���Ǝv���Ă��܂��B
|

�֓��@���q
�@
��t�������������� �w�|��
|
2013.02.03
No.9 �@
�@ �X�y�C���̎��R�ی�̈�[���_�Ԍ��� �Ɂ@���@���@�v
�ʐ^�́A�C�x���A�����i�X�y�C���j�̖k���吼�m�݂ɉ����ē�����300km�ɓn��̂т�J���^�u���A�R������Picos de
Europa���������̓��[�̋��E�̓��ɂ���n�C�C���O�}�̒����i�Ƃ��̂��ɗ����j�ł��B���̒����̓n�C�C���O�}�̕ی�‧���ƂɎ��g�ރJ���^�u
���A���i�����͂P�B�P���j�ɂ��ݒu���ꂽ���̂ł���Ƃ̂��Ƃł��B
�@���́A��N�i2012�N�j�X���ɃX�y�C���ŊJ�Â��ꂽ�u�v���X�~�h�����w���ۉ�c�v�ɎQ�����A�������ʂ̕����܂����B���̐܂Ƀs���l�[�R���t�߂ƃJ
���^�u���A�R���t�߂𗷍s���A�`���E�̍̏W�Ɗώ@������@��Ɍb�܂�܂����i�R�s���ɂ��ẮA���ĐM�B��w��w�@�Ńc�L�m���O�}�̐��Ԍ��������Ĕ��m
�̊w�ʂ����A���݃G�R�c�A�[�Ƃ���|���Ă���p���x�O�ݏZ�̃I�����_�l�Ɉē����Ă��炢�܂����j�B��L�̃n�C�C���O�}�̒����͂��̓r���ő����������̂�
���B����̗��s�́A���[���b�p�̃n�C�C���O�}�Ɋւ��钲���Ȃǂ̖ړI�ōs�����̂ł͑S������܂���ł������A���̋@��Ɉē����Ă��ꂽ�I�����_�l�̘b��C
���^�[�l�b�g�Œ��ׂ��X�y�C���̃n�C�C���O�}�̕ی�‧���Ƃɂ��ďЉ�����Ǝv���܂��B
�@���[���b�p�����̃n�C�C���O�}�͐�ł̊�@�ɒ��ʂ��Ă���A�t�����X�ƃX�y�C���̍����ɉ������s���l�[�R���ł́A1900�N�㏉���ɍŌ�̖쐶�̂�
�ˎE�����ł����Ƃ݂Ȃ���܂����B���̌�A���[���b�p�̑��n�悩��̎��Y�̂����x���ړ�����܂����B�������A���̒n��ŗL�̈���Ƃ݂Ȃ����̂���
���c���Ă����悤�ł����A�Ō�̌̂����N�O�Ɍ���ĎˎE����Ă��܂����Ƃ̂��Ƃł��i�ٔ������ɂȂ�܂������A�ˎE�����l�́u�g����邽�߁v�Ƃ̌�����
���F�߂�ꂽ�j�B�܂��A���̒n��͗r�⋍�̖q�{������ŁA�Ċ��͕��q���s���Ă��邽�߁A���Ȃ���ʔ�Q���o�Ă���A���Ƃ͊ȒP�ł͂Ȃ��悤�ł��B
�@�܂��A�X�y�C���̃J���^�u���A�R���̃n�C�C���O�}�ɂ��ẮA�J���^�u���A���ɐ�������M�������̓��ł͗B��n�C�C���O�}��1989�N�ȗ���Ŋ뜜��
�iendangered species�j�Ɏw�肳��A�ی�‧���Ƃ����{����Ă��܂��B
�@�X�y�C���̃n�C�C���O�}�̕ی�́A1973�N�Ƀn�C�C���O�}���܂މ���ނ��́u�쐶�����̕ی�v��ړI�Ƃ���@�������肳��A���̖@�Ɋ�Â���X�̋K��
�����ꂽ���Ƃ���n�܂�܂����B���̌�A1989�N�ɂ́u���R�n��Ɠ��A�����̕ۑS�v��ړI�Ƃ���@�������肳��A��Ŋ뜜��ɂ��Ă͉̂��߂̌v
������肷�邱�Ƃ����߂��܂����B�����ŁA���̖@���Ɋ�Â��Ă܂��n�C�C���O�}���܂ޑS���Ő�Ŋ뜜��J�^���O���Ҏ[����܂����B�ŋ߂ł́A2007�N
�Ɂu���R��Y�Ɛ������l���v�Ɋւ���@���ɂ��A���ʂȕی��K�v�Ƃ���쐶������̃��X�g�ƑS���Ő�Ŋ뜜��J�^���O���Ҏ[����A2008�N����̖@��
�ɂ��n�斈�̐�Ŋ뜜��J�^���O�Ҏ[�����߂��A�J���^�u���A���ł��n�C�C���O�}���܂ސ�Ŋ뜜��J�^���O���Ҏ[����܂����B
�@�J���^�u���A���ł́A1989�N�̖@���Ɋ�Â��Ē����Ɂu�n�C�C���O�}�v��v�����肳��܂����B���̌v��ł́A�J���^�u���A�R���S�̂̃n�C�C���O�}
�W�c�̈�`�I���l���Ǝ�Ƃ��Ă̐����������߂�̂ɕK�v�ȓ����̏W�c�Ԃ̌𗬂𑣐i���邱�ƂƂȂ�܂����B���̂��߁A�܂������̃R�A�ɂȂ�W�c�̕ی�A��
�B��}�邱�Ƃ�ړI�Ƃ��鎖�Ƃ��n�܂茻�݂Ɏ����Ă��܂��i�����͕ʂ̏B�A���ɑ����Ă��܂��j�B1989�N�̖@���ł́A�N�x���Ɏ��ƕƕK�v�Ȍv��ύX
��q�{•�_��•���Ƒ�b�i���A�q�{•����•�n��J����b�j�ɒ�o�����F�邱�ƂɂȂ��Ă��܂��B
�@2011�N�x�̕��i65�ŁA�J���[�j�ɂ́A���Y�N�x�̎�X�̃��j�^�����O�̃f�[�^�ƂƂ��ɂ���܂łɏW�ς��ꂽ�f�[�^�x�[�X�Ɋ�Â��܂Ƃ߂̐}�\
�i�n��ʂ̖ڌ����A�e�A��̎��O�}�̖ڌ����A�ƒ{�̔�Q�����̐��ڂȂǂȂǁj��ʐ^�i�����A�ߊl‧���b�̗l�q�Ȃǁj�������f�ڂ���Ă��܂��B�ō�30��
���[���i�I�j�̔����ƒ�߂��Ă���ɂ�������炸��������A�܂����̒n��ł��ƒ{��_�Y���ɑ����Q�͏��Ȃ��Ȃ��A��Q�ɑ���⏞���傫�Ȗ���
�Ȃ��Ă��܂��B�ʂ̎����ɂ��A�J���^�u���A�R���S�̂̃n�C�C���O�}�̌̐��͑�����140�����x�A�J���^�u���A���𒆐S�Ƃ��铌���̌Q��30���`
40���ɂ����Ȃ��Ƃ̂��Ƃł��B
�@�����m�̂悤�ɁA�X�y�C���͌��݂���߂Đ[���Ȍo�ϊ�@�Ɋׂ��Ă��܂��B���ł͂��̂��ƂɌ��y���āA�l�I�A���I�����̌����I�ȉ^�p�i�d���̉����
�ǁj�̕K�v���Ɛv���Ȕ�Q�⏞�̂��߂ɕی����x�𗘗p���邱�ƂȂǂ�����Ă��܂��B����ɂ��Ă��o�ϊ�@�̐[���ȃX�y�C���̈�n�������c�̂ł��ꂾ��
�̕ی�‧���Ƃ��p���A���{����Ă��邱�Ƃɂ͋�������܂����B�J���^�u���A���̓T���^�f�[���s�Ƃ����吼�m�݂̃��]�[�g�n�i�����A�M���̕ʑ��n���吼
�m��]�ދu�ɗ������сA�C�M���X�Ƃ̊Ԃɑ�^�D�ɂ�����t�F���[�q�H������j�����鏬���Ȓn��ł���A��r�I�o�Ϗ��u�܂��v�ł���Ƃ����w�i������
�����m��܂���B����ɁA�����Ȍ�Ɉ�x�͐X�т�O��I�Ƃ�������قǂ܂łɔj�����[���b�p�i���ɁA�����[���b�p�j�ł́A�i�ߖłڂ��I�Ɂj���ۑS�A
���R�ی삪���A�n�������c�̃��x���ł������ɍs���Ă���Ƃ����w�i�����肻���ł��B
�@���{�̖k�C���̃q�O�}�Ⓦ�k���璆���ɂ����Ẵc�L�m���O�}�ɂ��ẮA���Ȃ��Ƃ����ʂ͐�ł̊�@�ɂ͂Ȃ��A���R�̐H���A���̒��_�ɗ��N�}�Ɛl�Ƃ�
�����̂��߂ɕی�E�Ǘ����ׁX�Ȃ���s���Ɩ��Ԓc�̂Ȃǂ̋��͂̂��Ƃɐi�߂�����܂��B�������A��B�ł̓c�L�m���O�}�̊m���Ȗڌ��Ⴊ�₦�ċv��
���A�l���ł���ł̊�@�ɂ���܂��B�����̒n��ł͕ی�‧�v��̍����ڎw���đ��}�Ɍp���I�Ȍ̐������ƕی슈�������{����K�v������Ǝv����
���B
�@�Ƃ���ŁA�`���E�̍̏W�Ɋւ��ẮA�t�����X�ł͍��������A���R�ی�����Ȃǂ̊O�ł͋K��������܂��A�X�y�C���ł͖@���ɂ�肢���Ȃ��̃`���E��
�N�ł����Ă��i�X�y�C���l�ł����Ă��j�̏W���邱�Ƃ����̂��֎~����Ă��܂��B�X�y�C���l�̗F�l�i�����w�̌����ҁj�́A�����uNo one
cares.�v�ƌ����āA����������ǂ����Ƃ̈ӌ��ł����B�������A���p�̃g���u��������邽�߂ɁA�ނɎ�`���Ă�����č���̗��s�ŗ������\��̕���
�̌��ɌʂɃ`���E�̍̏W���\����O�����čs���܂����B�S���҂̍D�ӓI�ȑΉ��������āA����悭�R�����狖�邱�Ƃ��o���܂����B�s���l�[��J���^
�u���A�ŏo��������s�ҁA�n�C�J�[�A�r�����A�ѓ��Ǘ��҂̐l�������T���čD�ӓI�ŁA���l���̐l�����͎��̃��[���V�A�́i���R�j�`���E�̕��q�n���n���̌���
�̐����ɋ����������Ă���܂����B�����̐l���������S�ł������̂������ł������c�c..�B���Ăł̓t�B�[���h�Ń`���E��‧�̏W���Ă��ē��Ǝ҂ɏo��
�����Ƃ͂܂�����܂��A����������ł����B�����I�ȊW����X�y�C���̎R�Ŏq�������ɏo����Ƃ��Ȃ������̂͏����c�O�Ɏv���܂����B�u�`���E��߂�
�͈̂������Ƃ��v�Ƃ����悤�ȋ��炪�s���Ă��Ȃ���悢�Ǝv���܂��B
|

�ɓ��@���v
�M�B��w�@
���w�������Ȋw�ȁ@
���̐����w
���C�����@
|
2012.07.02
No.8 �@
�@ �����̖��͂�`����n���ȓw�͂��K�v�ł���B �h�@�q�@�d
�����̑��l���̎ړx�̈�͎퐔�ł���B����܂łɋL�ڂ���Ă��铮���̎퐔������ƁA�����͂��悻�V�����߂Ă���B������A�����͒n��������Ƃ����l
�����������Q�ł���A�n���͍����̘f���Ƃ������Ă���B������̍����ł��A�n��ɂ���Č`���������A�F�ʂ��قȂ����肵�Ĉق������邱�Ƃ��悭����B
���̂悤�ȑ��푽�l�������߂č����̏W�ɖ����ɂȂ鍩�����N�������B�������A�������N�̌����͊m���ɐi��ł���B���V�搶�A������Ɣ�������̃R����
�ł������Q���Ă���A���̌����Ɖ��P����q�ׂ��Ă���B�����������ΏۂƂ��Ă���l�ԂɂƂ��Ă͎₵������ł���B�������A����͎���̗���ł���
��A�����ȒP�ɂ��̗����ς��邱�Ƃ͓���Ǝv���B�ł��A�������N���������Ă��Ă��A�����̖��͂͌����Ă���킯�ł͂Ȃ��B����̂����鐶�����ɓK
�����A�l�X�Ȍ`�ԂƋ@�\�̑��l���𐋂��Ă���͍̂����ł���A�����̐i���E���l���Ɋւ��Ă͍��������ł͌�邱�Ƃ͂ł��Ȃ��B�n���ɍ����̖��͂𐢂̒���
�`���A�q�ǂ����҂������w�̌����Ɉ������邱�Ƃ��厖�ł���Ǝv���B
�@�����g���ŏ��Ɋ����������̖��͕͂ϑԂƋx���ł���B�𒎂Ɩђ������������X�ɂȂ�͉̂��Ƃ��_��I�ł���A�̂̑�ӂ��ŏ����ɗ}���A�H�ׂ����܂��Ɉ�
�������z�����Ƃ͔@���ɂ������������Ɖf��B������w�ō����x���@�\�̌����Ɍg����Ă����̂����˂Ă������������̖��͂������Ă�������ł��낤�B�I
�T���V�̌������n�߂�ƁA�����̌`�Ԃ̑��l���ɋ����䂩���悤�ɂȂ�A�����̏W���n�߂��B�q�ǂ��̍��A�������������Ƃ͂�����̂́A�����鍩�����N
�ɂ͉����y���A�I�T���V�͂������m��Ȃ������B��w�ł̌����͂����ς�g�D�A�זE�ADNA�Ƃ������~�N���̌����ŁA���������̂Ƃ��Ă������蒭
�߂����Ƃ��Ȃ������B�������A�I�T���V�̌����́A������Ƃ��̂��̂̓~�N���̐��E�ł͂�����̂́A���ʂƍl�@�͌́A�W�c�A��A�����͂���ȏ�̕��ތQ��
�ΏۂƂȂ�A�������̑S�̂�����悤�ɂȂ�A�����i���̌����ɑ��ݓ��ꂽ�B�I�T���V�̌������n�߂������̎��́A�n�����̌��������m�炸�Ɏ��s������d
�˂Ȃ���A�I�T���V�̌n���i���̌�������V�ȎO�搶�̂��ƂŐi�߂Ă������A�n�����̎}����������Ȃ���A�m���Ɍ`�Ԃ����Ă�����̂͌n������ɋ߂��W��
����A�t�͉����Ȃ邱�Ƃ�������ADNA�z��̏��ɐ����̐i���̗��j�����܂�Ă��邱�Ƃ��n�߂Ď����������͖{���Ɋ��������B�������A�S���͂����Ƃ͌�
�炸�A���炩�Ɍ`�����Ă���̂ɁA�n������ł͉����W�ɂȂ�A�t�ɖ����ɈقȂ�`�����Ă���̂ɁA�n������ł͎o���W�ɂȂ�A���������s�v�c�Ȍn����
��ꍇ�����X����B���̂��낤�B����̌n������`���ƁA�n��I�ɋ߂����̂��n������ł܂Ƃ܂邱�Ƃ͈�ʓI�ł���B�n��I�n�������̐������̂��A�܂�
���̒n��n���Ԃ̋��E���ǂ��ɂ���̂��A���������₢���o�Ă���B�v����ɁA�n�����������قNj^������������B�����������^����������ɂ͍X�Ȃ�
��͂��K�v�ł���A���R�ޗ��̒lj����W���K�v�ƂȂ�B���̂悤�Ɏ��̍����̏W�͕��q�n���̌�������n�܂����̂ł���B�̏W�͋�J���������A��J�����ߒ���
�o�ĕK�v�Ȃ��̂�����ꂽ�Ƃ��̖������͉����y�������̂ł���B�I�T���V�̌������n�߂Ĉȗ��A���N�t�B�[���h�ɏo�����āA���̂悤�ȍ̏W�̊y������
����Ă���B�����̏W���d�˂邱�Ƃɂ���āA�V���ȋ����[�����ۂ�������A�����̍X�Ȃ锭�W�ւƂȂ���B�����Ō��������̂́A�����̏W��ʂ��č�����
���͂����X�Ɋ�����邱�Ƃ�����A�����̂��������ō����̖��͂�m���Ă��獩���̏W���n�߁A���������ւƔ��W���邱�Ƃ�����Ƃ̂��Ƃł���B�q��������
���R�ɒ��ɐڂ���@��v�X���Ȃ��Ȃ�A���̎���ł͍����w�����҂̒n���ȓw�͂��v�X�d�v�ɂȂ��Ă���B
�@JT������������(BRH)�ł́A���N�̉ċx�ݒ��ɃT�}�[�X�N�[���Ƃ����C�x���g���s���Ă���B���̌������ł́A�g�߂ȍ������ޗ��ɂ��āA�Q���Ԃ�����
��DNA���o�APCR�ɂ��ړI��`�q�̑����A����z��A�n�����쐬�ƌ��ʔ��\�Ƃ�����A�̍�Ƃ��Q���҂ɑ̌����Ă�����Ă���B�u�����ڂɋ߂�����
���n������ɂ��߂��v�A�t�Ɂu�����ڂ��x����Ă͂����Ȃ��v�Ƃ�����|�̂��ƂŁA������̑I����s���ADNA�ɐ������̐i���̗��j�����܂�Ă��邱�Ƃ�
������Ɠ����ɁA�������@���ɕs�v�c�ŁA���͂̂��鐶�����̂ł��邱�Ƃ������ł���������Ă��炤�̂��ړI�ł���B�Q���҂ɂ͈ӊO�ɂ����w���������A����
�͍������N�����܂Ɍ����B�T�}�[�X�N�[�����Q���������ƁA�����ɋ����������A�����̏W���n�߂��q�ǂ�������B�����A�Q���҂̒����獩���w�����҂������
������������������Ƃ͂Ȃ��B
�@BRH�ł͂�����C�x���g��N�R��s���Ă���B����͎��������w�c�A�[�ł���B���ɂ̓c���O�������u���g���ēy�듮�����ώ@����v���O����������A�q
�������ɂ͏�ɐl�C�Ă���B���R�̗��t���̓y���̂��Ă��ēd�����g���ォ��M�������ēy���̏����������𗎂Ƃ��Č������Ŋώ@����B���̒��ł̈�ʓI
�펯�i�H�j�ł͌���ꂪ���̃��J�f�ނ�X�f�ނȂǂɑ��Ă��A�q�������͊��Ő^���Ɍ��Ă���l�q������ƁA���ݓI�ȍ������N�i�����j�͐̂������ς��
�����݂��Ă���Ɗ�����B���ɐڂ���@�������A�������������ݓI�ȍ������N�i�����j�͂����Ɩ{���̍������N�i�����j�ɂȂ��Ă����ƐM���Ă���B����
�������@����̒��ɑ�����A�₠�A��X�����̋@��𑝂₷�w�͂����Ȃ���Ȃ�Ȃ��̂��낤�B��������������N�̎��オ�܂��߂��Ă���̂����ł͂�
����������܂���B�����肢�����B
|

�h�@�q�d
�i�s�����������ف@
�n���i���������@��C������
|
2012.04.09
No.7 �@
�@ �f�����������̐V���㓞�� �V�@���@�P�@�K
|
�@���邳�烊���[�R�����̃o�g�����n���ꂽ�B���邳��Ƃ̍ŏ��̏o��́A2002�N�t�R�̐��Y�����ی𗬃Z���^�[�ōs��ꂽ�������[�N�V���b�v�̂�
���������Ǝv���B��ۓI�������̂́A���邳�p���������ޗ��̎a�V���ɂ���A����܂Ō������Ƃ̂Ȃ�������������J�Q���E�̈��́A���̋N���ɔ��邽��
�̊i�D�̍ޗ��ł������B���̌���l�X�ȋ@��ł��������铌�邳��̌����O���[�v���p���錤���ޗ��̃��j�[�N���ɂ́A�������₦�Ȃ��B�̏W�⎔��͍���Ȃ�
��������Ȃ����A�N�����������Ƃ̂Ȃ������������ł��邱�Ƃ͑f���炵���Ǝv���B�c��Ȏ퐔���ւ鍩�������������̑�햡�́A���ɑ��l���ɂ���Ǝv���B��
�̍����ɒ��ڂ���̂���������d�v�Ȍ��ƂȂ�B�Ⴆ�A�n����d�v�Ȉʒu�ɂ��鍩����f�������ł͖��炩�ɂ��邱�Ƃ̂ł��Ȃ������[�����ۂ���������
�ǂł���B����܂ł́A���Ƃ������[�����ۂ����݂��鍩���ł����Ă��ADNA�̌��t�ŗ������邱�Ƃ͍���ȏꍇ�����������A���̏͋}���ɂ�����
��B
�@1987
�N�ɔ��\���ꂽPCR�@�̕��y�ɂ��A�ǂ̐���������e�ՂɈ�`�q���������N���[�j���O�ł���悤�ɂȂ��ċv�����B���́A�ړI�Ƃ��錻�ۂ�������������
��`�q���ǂ�����Č����邩�ɂ���B�����̃Q�m�����ɑ��݂����`�q���́A�P��������x�ł���B���̗L���Ȉ�`�q�̒�����A�ړI�̈�`�q�������邽
�߂̃A�v���[�`�͗l�X�ł��邪�A�ǂ̕��@��p���Č����o�����͌����҂̘r�̌������ł���B�ŋ߂ł́A�R���s���[�^�[��ł̃N���[�j���O���\�ɂȂ����B
�S�Q�m�����̉�ǂ��s���v���W�F�N�g���i�s���Ă��邩�炾�B2000�N�̃L�C���V���E�W���E�o�G�̃Q�m����ǂ���ɁA�l�X�ȍ�����ŃQ�m����ǂ̕�
�������X�Ɣ��\�����悤�ɂȂ��Ă����B���̏�����ɉ���������e�N�m���W�[���o�ꂵ���B������V�[�P���T�[�ł���B1000�h���Ńq�g�̌l�Q�m��
����ǂ��鎞�オ�������Ȃ��A������V�[�P���T�[�̕��y�ɔ����A�{���̂��Ƃ��e�퍩���̃Q�m����͂��i�s���Ă���B�z��̃z�����W�[�Ɋ�Â��N���[
�j���O�̋�J���l����ƁA������1000�h���ō����̃Q�m���̊T�v����ǂł���Ƃ͊���������ł���B���M���ׂ��́Ai5k (5,000
Insect Genome
Project)�Ƃ���5,000��̍����̃Q�m����������5�N�Ԃʼn�ǂ���v���W�F�N�g�ł���B�����_�ł͂ǂ��܂Ō����������邩�͕s���ł��邪�A
�V���������Ɋ�Â����X����V�[�P���T�[�̊J�����i��ł��錻����ӂ݂�ƁA���̃v���W�F�N�g����������͉̂��������ł͂Ȃ��������B
�@
���݁A���f�������̑�\�͂����܂ł��Ȃ��L�C���V���E�W���E�o�G�ł���B���[�K���ɂ��1910�N�̔���ψّ̂̔����ȗ�100�N�ȏ�̗��j�����V���E
�W���E�o�G�����������́A�����̃��f���Ƃ�����萶���̊�{�������������������߂̃��f���Ƃ��Ĕ��W���Ă����B���f�������̒��̃��f���Ƃ��ĕs���̍����
�������闝�R�͑��X���邪�A��`�q�̋@�\��̓V�X�e���̖L�x���͈������B�����q�ɂ���`�q�g�����̂̍�o��1982�N�ɕ���Ĉȗ��A�l�X�ȉ��ǂ�V
�Z�p���������ꌻ�݂Ɏ����Ă���B����A�f�������̈�`�q�g�݊����̂̍�o��piggyBac�x�N�^�[�̊J���ɂ��傫���i�W�����B
1998�N�̃`�`���E�J�C�~�o�G�ł̐����ɑ����A���̂��Ƒo���ځA�����ځA�����ځA�����ځA�����ڂ̊e�퍩���ɂ����Ĉ�`�q�g�����̂̍�o������A��
���f�������ɂ����Ă���`�q�@�\��͋Z�p�����X�ɐi�W���Ă���B
�@
��`�q�̋@�\��͂ɂ�����v���́A1998�N�����Ŕ������ꂽRNA���iRNAi�j�ɂ���Ă����炳�ꂽ�Ƃ����Ă��ߌ��ł͂Ȃ��B��`�q�g�����̂̍�o
������ȍ����ł����Ă��ARNAi�@�ɂ��e�ՂɈ�`�q�̋@�\�j�Q���\�ɂȂ����B���̕��@�́A���S�ϑԍ�����s���S�ϑԍ��������łȂ����ϑԍ����̃V�~
�ɂ����Ă��L������������A�u���Ԃɕ��y�����B�������Ȃ���ARNAi�@�����\�ł͂Ȃ��͎̂c�O�ł���B��{��RNA��̍o�ɒ��˂������e��g�D�̍זE��
�Ɏ�荞�܂�Ȃ�����RNAi�������Ȃ��ꍇ��������ɂ���đ��݂���BRNAi�͖{���^�j�����̐����ɕK�{�̐��̖h��@�\�ł��邽�߁A�ǂ�ȍ����ł���
�E���ɓ�{��RNA�������Ă��܂���RNAi�͐�����B���ɁA�זE�����`�������ȑO�̑��j���E����̏�����ɓ�{���q�m�`�𒍎˂���ARNAi�ɂ��
��`�q�@�\�j�Q���ǂ�ȍ����ł�������ƍl������B���̂Ƃ���A������Ɉˑ�������{��RNA�̍זE���ւ̎�荞�݂̍��ق������郁�J�j�Y���͕s���ł�
��B���̌������𖾂����A�߂������ǂ̍����ł���`�q�̋@�\�j�Q���p�ӂɂȂ鎞�オ���邩������Ȃ��B
�@
�����ŋ߂̒��ڂ��ׂ��e�N�m���W�[��TALEN�i�s�������������������������@�`�����������������|�k�������@�d���������������@�m���������������j
�ƌĂ��l�H�k�N���A�[�[�ɂ��z����ٓI�Ȉ�`�q�^�[�Q�e�B���O�@�ł���B���̐�����́A�t�^�z�V�R�I���M�ƃJ�C�R��2012�N�̊w��ŕ��ꂽ�B
����A���̕��@�ɂ��f�������̈�`�q�@�\��͂��傢�ɐi�W���邱�Ƃ����҂����B
�@�V�����e�N�m���W�[�̊J���ɂ����@�_�I�ȏ�ǂ��ł��ӂ���A�u���[�N�X���[��
�Ȃ錤���ւ̔��W��������Ă����B���f�������Ɣf���������킯��Z�p�I�Ȋ_���͂܂��܂��Ⴍ�Ȃ��Ă���悤�Ɋ�����B����̍����c�m�`�����̔��W���y
���݂ł���B
|

�V���@�P�K
���É���w��w�@�����_�w�����ȁ@���������Ȋw��
����
|
2011.11.22
No.�U �@
�@ ��r�����w���番�q�n���n���̐��E�� ���@��@�K�@��
|
�@�u�����{��k�Ёv�̘b��i�i������j����u��`�q����̕W�{�Ǘ��p�v
�i�q������j�ցA�����Ď��ւƃR�����������[����Ă����B�����g�A�Ôg��Q�ɂ��n��̌Q��ł̉\���������Ƃ����A�Ƃ���N���������̈�`�q��͂�
�ւ�邱�ƂƂȂ����B
�@��
�̌̌Q�́A������ꌴ�q�͔��d���t�߂̂��߁A����c������s�\�Ȃ̂ł��邪�A�k�БO�ɕW�{���m�ۂ���Ă������߂ɁA���̂悤�Ȉ�`�q��͂̋@�����
�ꂽ�̂ł���B�ߗׂ̌̌Q����͒n���I�ɂ��Ȃ�u������Ă���A���������ł͂�����̌̌Q�ł��������ƁA���A�Ôg�̉e������قǂɊC�݂ɋ߂�
�ȂǁA���Ȃ���قȌ̌Q�ł��������炵���B����̐k�ЂƒÔg����X�̑z�������̂ł������Ƃ��Ă��A��N�E����N�Ɉ�x���炢�̃X�p���œx�X�N����
�Ă����悤�Ȏ��R���ۂł���ɂ͈Ⴂ�Ȃ��B�����ł���Ȃ�A�ߋ��ɂ��A�Ôg�ɂ��h���ɎN����Ȃ�����������Ă����̌Q�Ȃ̂ł́H�@�Ƃ����ӌ����ł�
���悤�B����������̏ꍇ�A�l�Ԋ����̉e���ɂ��̌Q�T�C�Y�̏k����������ꂽ��ɁA��K�͊h�����Ă���̂ŁA����܂ł̂��̂Ƃ͈Ӗ�������Ă���
�̂��낤�B
�@�����炭���k�n���̑����m���݂ɂ́A�����悤�ȋ����ɂ��鍩����Q�����ɂ�����
���̂Ǝv����B���̂悤�ȏ��ɂ����āA�u���̏ꏊ�ɁA���̍���������
�Ă����v�Ƃ��ĕW�{���㐢�Ɏc�����Ƃ�A��`�q���̂悤�ɕW�{����܂�����������āA������ƃA�[�J�C�u���Ă������Ƃ͏d�v�ł��낤�B���͂Ȃ���
���A���̂悤�Ȍ`�ł̍v�����ł���ƍl���Ă���B
�@�����ނ��܂ނ����鐶����Q�̈�`�q���́AGenBank��
��ы�����������Ă���f�[�^�x�[�X�ɏW�ς���Ă��邽�߁A�����ɓo�^���ꂽ�f�[�^�Ɋւ��Ă͖��l�����p�\�ƂȂ��Ă���B�����A���{�ɂ͐������Ă���
���悤�ȍ�����Q�̔z��f�[�^�Ȃǂ��d�ė��p�����Ă��������Ă���B�莝���̃f�[�^��GenBank��
��擾�����f�[�^�������Čn����͂��邱�Ƃ������̂����A���X�A�s���Ȍ��ʂ�����������邱�Ƃ�����B���̈���ɂ́A�o�^���ꂽ��`���ƁA���̈�`��
��̂�����ł������Ƃ��ēo�^����Ă���햼�̑Ή�����v���Ă��Ȃ����߂̂��Ƃ������悤�ł���B�ނ��A�ڂ�ȃ��x���Ƃ����������n����͂Ɏ���������
���錤���҂ɂƂ��ẮA�탌�x���̓��萸�x���������ʂ����E���邱�Ƃɂ͂Ȃ�Ȃ����߂ɁA����قNj����S�ɂ͂Ȃ�Ȃ��ł��낤���A�ł�����莯�ʂ̗e��
�Ȏ��ΏۂƂ���悤�Ȋϓ_����̃^�N�\���E�T���v�����O���Ȃ���Ă͂��邾�낤�B
�@�������Ȃ���A�����ΏۂƂ��Ă��鐅�������ނł́A��̑q������̃R�����ŏЉ�
���ꂽ�u�����ނ��v�̗�̂悤�ɁA������u���ʎ�v�Ə̂���Ă���悤��
��ɂ����Ă������B����̑��݂����炩�ƂȂ����肵�Ă���̂�����ł���B�b���ނ⒱�ނƈ���āA�}�C�i�[�ȍ�����ΏۂƂ��Ă��邪�̂̈ӌ��Ȃ̂�������
�Ȃ����A�q�����q�ׂĂ���悤�ȁu��`�q���o�ɗp����ꂽ�W�{�́A���ł����ފw�I�Č������\�ȏ�Ԃŕۑ������ׂ��ł���v���Ƃ�Ɋ����Ă���B
����ł��������X�y�[�X��肪���܂Ƃ����{�̔����َ�����l����ƁA�����I�ɂ͓�����ƂȂ̂����m��Ȃ����A�핪������舵����������L�ڂɊ֘A����
�����ɂ����āA��`�q��p����ꂽ�ꍇ�Ȃǂɂ����Ă͓��ɁA���҂̐Ӗ��Ƃ��Ĉ�`�q���o�ɗp�����W�{�ނ�R��ׂ��@�ւɎ��߁A�W�{�̏��݂�_�����ɂ�
���L���ė~�������̂ł���B�������邱�Ƃ�����ꍇ�ł��A���߂āA�̏W������Y�n��_�����ɏڂ����L����Ă���Č������Ȃ����ۂɂ͖��ɗ���
�ł͂Ȃ����낤���B
�@�����܂ŁA�k�Ђ̂��Ƃ��`�q��͂ɋ����ꂽ�W�{�̈����Ɋւ��鎄��
���q�ׂĂ������A�㔼�ł́A�������g�̍���DNA�����ɂ��ďЉ���Ă������������B
�@���̍��������̂͂��܂�́A�u��r�����w�v�Ə̂���镪��ŁA�����̑̂Â����
�ʂ��āA���l�ȍ����`�Ԃ̊�{�v�����𖾂炩�ɂ������A�Ƃ����Ƃ���ɂ���
���B�Ȃ��ł��A�������ł����n�I�ȍ����ނł���J�Q���E�ނɒ��ڂ��A���������Ђ�����`�Ԋw�I�ɁA�����đg�D�w�I�ɒNj����Ă����B�w�ʎ擾�܂ł�
DNA�Ƃ͖����ɉ߂����Ă������̂́A������`�w�I�������A�V���E�W���E�o�G���͂��߁A�J�C�R�K��R�N�k�X�g���h�L�ȂǂƂ��������f�������ނ���A�n���i
���̌���
����L�[�^�N�T�ւƏ��X�Ɋg����͂��߂����ł����������߁A�L�������ނ̐i���̌�������J�Q���E�ނł�������`�w�I�����ɊS�������͂��߂�悤�ɂȂ�
���B�������A���̎�̌�����@�ɂ͑S���̑f�l�ł��������߁A�l�X�ȕ��X�Ƃ̋���������ʂ��āA�J�Q���E�ނɂ����锭����`�w�I�Ȍ����ɂ�����˂����ނ���
�ƂȂ����B
�@���̂悤�Ȓ��A7�N���قǑO�ɁA���݂̐M�B��w���w�������Ȋw�Ȃɒ��C��
���B
�ꂩ�猤�����𗧂��グ�Ȃ���Ȃ�Ȃ��ɂ��������A��`�q��͂Ɋւ��ẮA���Ȃ�̕������w�ȋ��ʋ@��ނ𗘗p���邱�ƂŃJ�o�[�ł������߁A�T�[�}
���T�C�N���[�≓�S�@�A�①�E�Ⓚ�ɂ₻�̑��̏����ނ��������Ă��܂��Β����Ɍ������X�^�[�g��
���闘�_���������B�����āA�L���Ȏ��R�����͔����ɑ��݂���M�B�̒n�̗��͉��������肪�����A����܂ł͑S���o���̂Ȃ��������q�n���n���w�I�Ȍ����ɂ�
�V�K���肷�邱�Ƃ������B����ɍK�^�������̂́A��w���Ɂu�R�x�Ȋw�����������v�Ƃ����g�D�������オ��A���̌������̌��C�ɂ��Ă������������Ƃł�
��B���{�A���v�X�n��𒆐S�ɁA�R�x�Ȋw�ɊW����l�X�ȏ�W�ς��A�t�B�[���h�ł̒����E�����ɂ��X��}���Ă���������Ƃ����A���̏�Ȃ�������
�������Đ������̂ł���B
�@
���ɒ��肵���̂́A��l�I�O���i��180
-100���N�O�j�̑��䍂�A��̎R�x�`���i���N�j�ɂ���ĉ���������ɂȂ����Ǝv���錴�n�I�J�Q���E��Q�Ɋւ��錤���ł������B���ۂɈ�`�q��͂�����
�݂��Ƃ�
��A�����ɑ��䍂�A��̓����i���{���Ɣ�ˑ��j�ň�`�I�����������Ă��邱�Ƃ����炩�ƂȂ����B�����āA���肳�ꂽ���R�[�̌̌Q�Ԃł̕���N����n�j��
�悭���v�����
�̂ł������B�܂��A���̎R��ł́A�Ċx�ΎR�Q�̊������ɔ����A�͐�̍ǎ~�߂Ɨ��H�ύX���p�ɂɐ����Ă����Ƃ�����邪�A���̂悤�ȁu�͐쑈�D�v���ۂɊւ�
�Ă��A��`�q��͂̌��ʂ͋����[���֘A���������Ă��ꂽ�B
�@���̂悤�ȓW�J�����������ɁA�R�x��ɂ����镪�U�͂̎ア������Q�ɒ��ڂ��邱
�Ƃ͓��P���Ȃ�����A��͑ΏۂƂ��鍩���Q���g���Ă����B�ŋ߂ł́A�Ώۂ�
�n����g���āA�쐼�������܂߂����{�L��A����ɂ͒��N������嗤�Ȃǂɂ������^�Ԃ悤�ɂȂ��Ă����B���̂悤�Ȍ������͂��߂������A���݂̕��z����
�ʂ������̂ł���A�����悤�Ȑi���j��H���Ă����̂��낤�ƍl���Ă����̂����A���ۂɏڂ�����͂����Ă݂�ƁA�����P���Ȃ��Ƃ���ł͂Ȃ����Ƃ��悭
�������Ă����B�ЂƂ��Ɂu���{�̍����̋N���⍩�����̐����v���Z�X�v�Ƃ����Ă��A���ɑ��l�ȃp�^�[����������̂��ƁA�����A�y���݁A�����ĕs�v�c����
��Ɋ����Ȃ���߂����Ă���B
|

����@�K��
�M�B��w���w�������w��
����
�i���C�@�M�B��w�R�x�Ȋw�����������j
|
2011.10.24
No.5 �@
�@ ��`�q����̕W�{�Ǘ��p �q�@���@�ǁ@��
|
�@�w��
���ނ��x�Ƃ��鍩��������B�w�����ނ��x�́A���������Ɨ�����̐��ɐ��ސ��������i�c���j�̑��̂ŁA�M�B�Łw�����ނ��x�͗L���ȐH�ނł���B�w����
�ނ��̒ώρx�Ɏg���l�X�ɂ��Ȃ��݂̐[���q�Q�i�K�J���g�r�P���̈�`�q���e�n�ō̏W���Ē��ׂĂ݂�ƁA����܂Ńq�Q�i�K�J���g�r�P���Ɠ��肳��Ă�����
�̂̒��ɕ����̎킪�������Ă������Ƃ����炩�ƂȂ����B���������q�Q�i�K�J���g�r�P���̋߉���́A�q�Q�i�K�J���g�r�P�������܂�ɕ��ʎ�Ȍ̂Ɍ���
����Ă����̂�������Ȃ��B
�@��`�q�Ŏ��ʂ��ꂽ������c������A��̔F���ɕK�v�ȊO���`�Ԃ̏�W�܂�A
���{�̉͐쐶�Ԍn���\����q�Q�i�K�J���g�r�P���̋߉��킪�V�����^�N�T��
���Ēlj�����悤�Ƃ��Ă���B������`�q����͂����̂��ǂ̌̂�������Ȃ��悤�ȏ�ԁA���Ȃ킿�W�{�̐������[���ɂ���Ă��Ȃ������Ȃ�A���̌�����
�����ł��Ȃ������ɂ������Ȃ��B��`�q��͂��ꂽ�̂�����ł��āA������`�ԉ�͂ŗ��ł��ĐV�����^�N�T���F���ł����̂ł���B
�@��`�q���i����z��j�́A���ތQ���i�����̊w���j�ƌ��т����Ă���B����
��`�q���͂��������̓���Ɍ�肪����A���͂����������ǂ̌̂���
��ł������ؕs�\�ȏꍇ�A���̏���p�������ʂ̉��߂ɑ傫�ȉe�����o��͎̂����̗��ł���B���̈Ӗ��ň�`�q��͏؋��W�{�́A���ފw�ɂ�����w�^�C�v
�W�{�x���Ȃ킿��̒�`�Ɏg��ꂽ�W�{�ɏ����鈵������ׂ��ł���B
�@��`�q��͂��ꂽ�����W�{�́A�ǂ̂悤�ɕۑ��Ǘ�����Ă���ł��낤���H��ʂ�
��`�q��͂Ɏg����W�{�́A�A���R�[���t�Z�W�{���������܂��܂Ȍ`��̕r
�ɓ����Ă��邱�Ƃ������B����䂦�i�z�˂���j�Ǘ�����ϔώG�ł���B�W�{�̕ۑ��ɂ��Č����̌���͑�ϋ�J�����Ă���B��J�̍ő�̌����͎��[�X
�y�[�X���[���m�ۂ���Ȃ����Ƃɂ����̂��B�i�����W�{�́A���x�ɋK�i�����ꂽ�w�h�C�c���x�Ƃ��錘�S�Ȗؐ��̔��Ɏ��[����t�Z�W�{�Ɋr�ׂĊǗ�����
�₷���j�B���x���ɏ������ޏ��͂��Ƃ��A���x���̍ގ���L���Ɏg���C���N�Ȃǂ̑I�����ێ��Ǘ��ɂ͏d�v�ȗv�f�ł���B
�@��`�q���o�Ɏg�����W�{�͂�
�ł����ފw�I�Č������\�ȏ�Ԃŕۑ������K�v������B�W�{�̈ێ��Ǘ��́A�����Ė����łł���悤�Ȃ��̂ł͂Ȃ��l�����
�܂߂��o��K�v�ł���B�����̊����W�{�̏ꍇ�A�����ق��l����W�{��a����A�W�{���ɔz��E�f�[�^�x�[�X�ւ̓o�^�E���J�ɂ�����o����Օi�Ɛl�����
�܂߂ĂP�̂������T�h���Ƃ��Ă��镶�������������A�t�Z�̈�`�q��͏؋��W�{�̏ꍇ�K�v�Ȍo���1�̂�����1500�~������Ȃ��ł��낤�B����͎�
���������Ă��鐅�������̏ꍇ�ł���A���ׂĂ̍����ł��ꂾ��������ƌ����̂ł͂Ȃ��B���\�o���������̂ł��邪�����̍̏W�⓯��ɂ����������Ԃ�J
�́A��`�q��͂ɂ�����
���o��A�����̌��؉\����S�ۂ���Ƃ������Ƃ����킹��Δ[���̂ł��Ȃ����z�ł͂Ȃ��B
�@�Ő�[�̂��Ƃ���ɖڂ������Ă���ƁA�v��
�ʏ��ɗ��Ƃ���������ł���B�������J�����đf���������������Ă��A�����؋��W�{���Ȃ��ƐM�p����Ȃ�����
�ł���B��`�q��͏؋��W�{�̖��́A���R�������������n�Ŕ����ق𗈊َҐ��ł����]���ł��Ȃ��悤�Ȍ����邩��S�z�ɂȂ�̂ł���B�W�{���������_
��Z�p�͊m���ɌÓT�I�Ȃ��̂ł͂��邪�A�Ő�[�̂������낢���������̂悤�Ȋ�b�̏�ɐ��藧���Ă��邱�Ƃ����̋@��Ɏv���N�����Ă���������K���ł�
��B
|

�q���@�Lj�
��t��������������
��Ȍ�����
|
2011.9.22
No.4 �@
�@�@�����{��k�Ђ���w�Ԃ��� �i�@���@�Á@�V
2011
�N8��7���A�����m���݂𑖂��鍇�Ԃ�D���āA�R�`�����̎R�ŕ�����T�������Ă����B30�x���鏋�����ŁA�G�]�[�~�̑升���̃X�M�т�������ʂ��A
�������\�N���ʂ�l���Ȃ��܂܂Ɏ���ꂽ�������x�������Ȃ���A�悤�₭������̌E�n���݂��ĉ����Ă݂��B�ۂނ����̊Ԃ��琁���o����C�ɁA�����}��
��₳��ĐS�n�悢�B�����ɂ́A����Ă͂�����̂̌����ȐΑg�݂��c���Ă����B���āA�����͋ߋ���~�̃J�C�R�̗��̒����ɂ������B�₽���̏�ɐQ�]��
�Ȃ���A�X�M�̏��𗬂��_�̍s����ǂ��Ă����B
���̉ẮA�Â�����̗{�\�𑱂���
�_�Ƃɗ��݂���ŁA�����Ԋu�ŁA�d���̗l�q���B�e�����Ă��������Ă����B94��70��
��l�̒j��ɂ�鉊�V���̃N������A�\���t��H�ׂ鉹�B�R�`���암�ł́A���łɗ{�\�_�Ƃ͂��̂P�������ɂȂ��Ă����B���̉Ƃ������A���ĕ����ɗV�т�
�s���ĐΎ��̒�������@�艺���A�X
�̉���r�߂ėV�Ƃ����b�����Ă����������B��C�̕����o�����ł͈�N���C�������肵�A�J�C�R�̉H�������߂ł���B�����̐Ύ��́A�l�Ԃ��C����
�Ǘ��ł��Ȃ���������́A�����̒m�b�̌����ł��������B
�①�ɂƂ������̂��o�ꂵ�Ă���A
������K�i���Ƃȁj���l���Ȃ��Ȃ�A�����I�̊ԂɐΑg�͕��ꂩ���āA�N���d����}�u�h�E
�̖��ɕ����Ă����B
3��11���̓����{
��k�Ђ̂��ƁA���R�j�����̔�Q�͂����Θb��ɏ�����B���O���c�̔����ق̕W�{�����ɑ����̐l�X���ւ�������Ƃ͗l�X�Ȕ}�̂ŕ�ꂽ���߁A�l�X�̋L
���ɐV�������낤�B
�c�m�`�����̗̈�ł́A�ЊQ���̎����ۊǂ͂ǂ����낤���B�ቷ�ŕۊǂ�
�˂Ȃ�Ȃ����̂��啔�����B��K�͒�d�Ƃ�����肪������ƁA�①�{�݂��@�\���Ȃ��Ȃ�B�c��ȗⓀ�������A�S���e�n�ɎU��镗���ɒS���グ�A�Ύ���
�[�����Ē�������킯�ɂ͂����Ȃ��B
�@
�܂��́A�a�@���w���̂悤�ɁA���p�d���ւ̐�ւ����\�Ȉ��肵���ۊǎ{�݂̊m�ۂ��O��ɂȂ�B�V�[�P���T�[���͂��߂Ƃ��������{�݂⎎��́A����
�̒��K�v�ɂȂ��ʂ����邩������Ȃ��B�Ɩ��p�Ⓚ�ɂ��������Ƃ��Ă��A���p�d�����S���ƂȂ��{�݂ł́A�ĂȂ�ΐ����̒�d�ł��n�����Ă��܂��B��
��ɁA��K�͂ȉЂȂǂ������₷���H�ƒn�сA���邢�͔R�������{�݂⌴�q�͎{�݂̎��͂Ȃǂ��A�ЊQ���ɉ����N���肤�邩�Ƃ������Ƃ�O���ɒu���Ă�����
�Ȃ�Ȃ����낤�B
�k
�В���ɁA���炭�g�����̎��Ȃ��������}������߂�Ƃ��A�����͎��͂̎d���̎ז��������Ă��Ȃ������A�����̖@��w�����ɍv���ł����ʂ͂Ȃ���
�̂��ƁA�Ȃ��m�b���i�낤�Ƃ����B�A�������ׂēr�₦���Ȃ��ŁA�c�m�`��͂̎���𓌋�����R�`�ɉ^�Ԃ��Ƃ͂ł��Ȃ����B�c�m�`��͂ɂ��l���ʂ̎��v
���}�����������Ƃ̎v�����������B���v���A�{���ɕK�v�Ȃ��͎̂��q���ً̋}�A���Ԃł����ɔ��������̂����A�����͂����܂œ������Ȃ������B�������A
����̉^������������Ȃ������B�Ⓚ�ł̑�z�ւ��ғ����Ă��Ȃ���ԂŁA�������̃h���C�A�C�X�͔�Вn�̈��u���ɑ����A���Ԃɉ���̂͂Ȃ���������
���B���ǁA���͋����̏�i�̕��X�ɐS�z�������������ŁA���i���ɗ����Ȃ��l�Ԃ͔�펞�ɂ����ɗ����Ȃ��A�Ƃ������Ƃ��ؖ����������ɏI������B
��
��������ʂł̊�@�Ǘ����A��������Ƃł��Ă��邱�Ƃ����Â����S�����̂��V���Ђ������B�H��̑傫�ȗ֓]�@�́A�d���̑���Ȃ����p�d���ł͓������A
�R�`�V���Ђł͏H�c�E�V���E�����Ȃǂ̊e�ЂƘA�����Ƃ�A���ʓI�ɐV���ň�����ċ}�����A�����̔z�B�ɊԂɍ��킹���Ƃ����B��d�������ăe���r�E�p�\�R��
�̎g�p���������Ԃł́A�W�ʂ̗Վ��łƂ͂����A�V�����l�X�ɉ^�傫�Ȏʐ^�̏��͏d�������B
�����ɒm�b����g���ė����̒������Ԃɍ��킹�����A�Ƃ����b���A�R�`�s
���̈��݉��ŎR�`�V���Ђ̃f�X�N���畷���Ȃ���r�[���̃W���b�L���X���Ă����̂́A4��7���̐[��̂��Ƃ������B���̒���A�ő�̗]�k���N�����ēX�̂Ȃ����^��
�ÂɂȂ�A�����Ɉ��͂��̃f�X�N�̕��X���u��d�ŗ֓]�@���~�܂�B���Ԃ��x������A�O��i3��11���j�����܂������I�v�ƌ��t�����킵�Ȃ���{�ЂւƋ}���ꂽ�B������
������8��30���A�V���ň�����ꂽ4�ʂ����̒���
���R�`�����̊e�˂ɓ͂���ꂽ�̂��B
���̐k�Ђ̂Ȃ��ɂ����āA���O�̖��S�ȑ�ɂ���Č�������������V��
�Ђ́A��@�Ǘ����ł��i��ł����g�D�ɈႢ�Ȃ��̂����A�@�ւ����Ƀj���[�X�ɂ��L���ɂ��o���A�ӊO�Ȃقǐl�X�ɒm���Ă��Ȃ��B
�@
��Ў҂͊F�A�u���ꂽ����A���ʂƂ��ċ��x�Ȑ��_�͂ŏ��邱�Ƃ��������A����ꂽ�W�{�̂��Ƃ��Ă��\���������Ɂu�ЊQ�ł�����v�ƌ����
��B�Ί���₳���A�����Ďx���ɑ��銴�ӂ������ɂ��Ȃ��B�ނ�̌�����{�����o�邱�Ƃ́A�����͂Ȃ����낤�B�����炱���A�u�x�������v
�u�������ꂽ�v�Ƃ����A�O���̎�������̔��k�ŏI����Ă��܂������ɂȂ�B
�Ƃ���ŁA�~���̐����オ�������I�����ق̕W�{�ɂ͑Ƃ�ꂽ���A�����ɏ���
����ʌl�����̉���A���̕W�{�A�ʐ^�A�m�[�g�A�������A�ƂƂƂ��ɊC��
���������ƂɒN���G�ꂸ�A�����ًƊE�̑g�D�I�x�����������グ���邱�ƂɁA���ǂ������������邱�Ƃ��m�����B���{�̎��R�j�����́A�قڌl�����̒~��
�ɂ���Ďx�����Ă������Ƃ͘_��҂��Ȃ��B�����ق̕W�{���R��B����̂悤�Ȕ�펞�ɁA�����������Ԃ̌l�����́A�c�O�Ȃ��犮�S�ɉᒠ�̊O�ɒu�����
�����B�ǂ̂悤�Ȕ�Q���������̂��Ƃ������̈ꌳ�������Ȃ���Ă��Ȃ����Ƃ́A���̓��{�̎Љ�̑傫�ȗ��Ƃ����ł�����B
�Ⴆ�A�����c�m�`������ŁA�Ⓚ���ꂽ�c�m�`�����̕ۊǏƁA�ЊQ���ɑz�肳
���댯���ɂ��ď����Ƃ�܂Ƃ߁A���ꂼ��̎{�ݓ��ł̑�Ɍ��т�
�Ă͂ǂ����낤�B�ۑ�̋��L�Ȃ����ẮA����������悤�Ȃ��Ƃ��J��Ԃ����B�������A�����ق̎x���������̓w�͂ɂ���Ďx�����Ă��邱�Ƃ́A��[��
�ւ�������̂Ƃ��Ă悭���m���Ă���B�����A���ꂾ������k�ɏI��点�Ă��܂��ẮA�W�{��m�[�g�ƂƂ��ɏ������ݖ�̌����҂̏�M���A���܂ł�������Ȃ��̂��B
|

�i���@�ÔV
���R�ʐ^��
|
2011.8.10
No.3 �@
�@�@��s�����i�K�T�L�A�Q�n����������� ���@���@���@��
|
�g�߂Ȑ��������̏W���Ė��O�ׁA��������������ώ@���邱�Ƃ���
�����w�͎n�܂�|�|�������̃x�r�[�u�[�}�[����́A������������߂Đ^�����ȉȊw������Ĉ�����B�����̏W�����コ��A�W�߂��W�{�����̂܂�
�Q����A���ꂪ�ċx�݂̏h��ɂȂ�B���R�����̃e�[�}�ɔY�܂���邱�ƂȂǂȂ��A�������N�ɂ͂��肪��������B�������̂ł���q���A�����������̂�
�Ă����̕����A�q�������ł̏���͂����ƍ��������B�����琢�̒��͕ς���Ă��܂����̂��c�c�B
�������N�̐��͌��݂̔�ł͂Ȃ��A�`���E�𒆐S�ɁA���Ɛ��A���邢�͓�Ɩk�̎q��
���ʂ�ʂ��čs���W�{�̌������͂�����B�G���u�q���̉Ȋw�v�������낤���A��]�̎�ƒł������L���ĕ��ʑ�������߂�R�[�i�[���������B
��������A���̃l�b�g���[�N�ɉ�������B�֓��A�����̎q��������
������A�����A�썑�E�F�{�s�ɏZ��ł������ɂ͂��Ȃ�L���������B��̒��ɂ̓~�J�h�A�Q�n�A�N���R�����T�L�A�T�c�}�V�W�~�ȂLj�H�v���Ȃ��Ɠ�����
�����̂����������A�u��͏��i�v�̃i�K�T�L�A�Q�n��c�}�O���q���E�����͎���̒�ł�����ł��̂ꂽ�B���ꂪ��B�ł͐�Ό����Ȃ��M�t�`���E��L�x���^
�e�n�ɉ����Ė߂��Ă���̂�����A����͂����~�߂��Ȃ��B���w�𑲋Ƃ��邱��̕W�{���ɂ́A���{�̓y����Ƃ���Ă����`���E200��̂W��
�ȏオ���Ԃ悤�ɂȂ��Ă����B
���ꂩ����I�A���̎��͋�B����͐�`�����ꂽ���l�s�ɋ����\���Ă���B10
�N�قǑO����A���Ƀi�K�T�L�A�Q�n��c�}�O���q���E�������p�������n
�߁A�嗤����̐l�דI�A�����Z���ȃA�J�{�V�S�}�_���ƕ���ŁA�����Ƃ����ԂɌ�������p�x�̍ł������`���E�ɂȂ����B����A���̎���̃`���E���͋�B��
��炵�����N���セ�̂��́A�����ɏZ�݂�������ɂ͑z�������Ȃ��������i�ł���B���\�N�������Ă͂��ƖK�˂Ă��Ă��ꂽ���J�m�Ƃ����C�����Ȃ���
���Ȃ����A��͂��a���̕����͂邩�ɋ����B�W�{�̕��X�����Ƃ��������������܂ő����Ă����Ƃ��Ă��A���݂̋�B�̎q�����ɂ͎��̂悤�ȘB���p���g���Ȃ�
�Ȃ��Ă��邱�Ƃ́A���Ȃ��Ƃ��m���ł���B
�`���E�Ɍ��炸���܂��܂ȍ����̖k�オ�b��ɂ̂ڂ�B�����ɂ��Ă͂�
���c�_���c��ɂ��Ă��A���{�̉��g���X���͂��͂⓮����������̂悤�Ɏ��ɂ͌�����B�i�K�T�L�A�Q�n�̗c�����H�ׂ�~�J����J���^�`�A�c�}�O���q��
�E�����̗c�����D�ރp���W�[�́A�֓��n���̎s�X�n�ɂ�������ł�����B�ނ�̕��z�g���j��ł������̂��~�̊����������̂ł���A�C�����オ��Ζk�i��
��͓̂��R�̂Ȃ�䂫���Ƃ������Ƃł���B
�ꌩ�A���͂̎��R�����₩�ɂȂ����悤�ɂ݂��邪�A���Ԃ��ł͂�
���B����n�̐��������ꂾ���̑��x�Ői�o���Ă���Ƃ������Ƃ́A�k���n�̐����͓��������̃y�[�X�Ō�ނ��Ă���Ƃ������Ƃ��B�L�����́A�����P�ɁA�V����
����悤�ɂȂ������͖̂ڂɂ��₷�����A���Ȃ��Ȃ������̂͋C�Â��ɂ����Ƃ����l�ԐS���̗��Ƃ����ɉ߂��Ȃ��B�����A�ߋ��̑����̐�ł��A�قƂ�ǒN��
���C�Â���Ȃ��܂܋N���Ă����B���Ӑ[���Ď����Ă��Ȃ��ƁA�Ƃ�ł��Ȃ����ԂɂȂ��Ă��܂����˂Ȃ��\��������B
�����Ƃ��A�P���Ȑ}���ɂ͓��Ă͂܂�Ȃ��A���g���̔����Ƃ��v���錻��
������������B���Ƃ��A�E�X�o�V���`���E�̐��͊g��B�{���͊���n�̃`���E�ł���͂����A�C���㏸�ƂƂ��ɕ��z���ɍL������A��n�ɉ��肽��A����
�`���E�Ƃ͋t�s���Ă���B�C��ω��ɂ����̂ł͂Ȃ��A�c���̐H�������T�L�P�}����������r�n���L���������Ƃ��������Ƃ�������L�͎�����Ă���悤��
���A�L���͈͂ɓ����悤�ȌX�������邱�Ƃ��炷��ƁA���ꂾ���������Ƃ��v���Ȃ��B�k�J�̃`���E�Ƃ��ėL���ȃX�M�^�j�����V�W�~�̕��z�́A�Ȃ�����k�J�n
��ɍL����n�߂Ă���B������́A�{���̐H���Ƃ����g�`�m�L�̔z��ւ��i�o���Ă��āA�H���̓]�����Ă���͖̂��炩���B
���g�����l�ׂ̂����炵�����̂ł���Ȃ�A����͂������čD�܂�������
�Ƃ͂����Ȃ��̂��낤���A������}���Ɏ~�߂�肾�Ă�����l�ނɂȂ����Ƃ��m���ł���B�����̕��z����C�ɏ�����������悤�ȋC��ϓ����킪�g�Ŏ���
�ł���悤�ȋ@�������������Ƃ͎v���Ȃ��BDNA���̂��̂̕ψق�A�G�s�W�F�l�e�B�b�N�ȕω��ȂǕ��q���x���̃`�F���W
�����łɎn�܂��Ă��邩�����ꂸ�A��������������Ɛ����̑��ݍ�p��m�邽�߂̑s��Ȏ����ƂƂ炦�Ă͂ǂ��Ȃ̂��낤�B���ԂƂ��Ă̐����̐i���⑽�l��
�ɔ���`�����X�����������̂��A�Ƃ������z�������Ă����̂��Ǝv���B
|

�����@����
�T�C�G���X���C�^�[
|
2011.6.20
�@No.2 �@
�@�@�u�̏W��������ցv �V�����������N�̈�Ă��� ���@�@�F�@�i
|
���͏��a�Q�W�N��
�܂�ŁA���S�t
��
�����ɂ͍��x�o�ϐ������n�܂��Ă���A���s�s���̏��w�Z���ӂɂ������n�A���c�A�т��N�X�Z��n�ɕς���čs���̂�ڂ̓�����ɂ����B����ł����w����
�ċx�݂͖����U�����璎���ŁA���t���O�̐��ⓔ�ɑO������J�u�g���V��N�����������ɍs�������������B���w�Z�̂T�A�U�N�̓`���E�ɋ������o�āA
���s�s���̃`���E��S��ޏW�߂Ă�낤�Ə��w���Ȃ�Ɏv���Ă����B���������w���ɂȂ�Ə�ɂ͏オ���邱�Ƃ�m��A�ނ̕W�{���ɖ�O�Ō������Ƃ��Ȃ��E�X
�o�V���`���E�������Ă���̂����ċ������B�e��Z�ɍ̏W�ɘA��Ă����Ă��炤�̂͂��邢����Ǝv�����B���w�Z�Ɠ����~�n�ɂ��������u�Б�w�̍����W�{�W
�����w�Ղ̍ۂɌ��ɍs���A�q�T�}�c�~�h���V�W�~�̕W�{�������Ă�������B�������ʂ�V�T�C���͂��̎����߂ċ����
���B�قǂȂ��q�T�}�c�~�h���V�W�~�̐����j���𖾂��ꂽ�Ƃ̋L�����V���Ɍf�ڂ���A���̃`���E������Ȃɒ��������̂ŁA����ɂ��ꂪ���s�ɂ��邱�Ƃ�m��
�ċ������B���s�k�R�̐������q�T�}�c�~�h���V�W�~�̐��n���Ƃ́A���w���̎��͒m��R���Ȃ������B�����T�`�P�O�Ώ�̐���͂���܂Ŗ��m���������{��
�`���E�̐��Ԃ����X�Ɖ𖾂��ꂽ����������߂����ꂽ�Ǝv���B��ʂ��s�ւŖ��J��ȏꏊ�����{�ɂ�������c���Ă���A�����ł̍̏W�Ɗώ@�ɂ���ē���ꂽ
�V�����m���͎R�̂悤�ɂ������B
�@��
�݂͂ǂ����낤�B�������N�͂�
�Ȃ��Ȃ��ċv�������A��w�̗��w�������Ȋw�Ȃɍ������D���œ��w����w���ȂLj�l�����Ȃ��B�g�̉��̍����͋ɒ[�ɏ��Ȃ��Ȃ苻���̑Ώۂ��̂��̂��g�߂�
���Ȃ��Ȃ����B�n�����D��┎���ق������̏W����J�Â��Ă��A�ȒP�ɂ͐V���Ȓm���������Ȃ��Ȃ����������̏W����Ӌ`�����̊y�������q���B�ɓ`���邱��
���ł��Ȃ��B��������̑�l�����́A�����������y���v���o�����̂܂܈��������āA�����悤�Ɏq���B�������������Ƃ��Ă���B�q���B���g�̉��̍�����
�̏W���ċ^��Ɏv�����Ƃ̂قƂ�ǂ͂��łɑ�l�B�����炩�ɂ��Ă��܂����B����ł͍������N�͈炽�Ȃ��B�������N���痝�ȍD�������܂�A�Ȋw�҂����܂��
�Ƃ������͂������݂��Ȃ��悤�Ɏv����B
�@��
�͉�������悢���낤���B��
�����̏W���`�Ԃ̍ו��ׂĕ��ނ�����A�n��ɂ���Ă킸���ɈقȂ����̈Ⴂ�ׂ���A���ǂ��ɉ��������������ׂ��肷��悤�ȁA�]�O�̍���
�����͖ʔ����Ȃ��̂ł���B���͎������Ǝv���B�q���͗��Ȏ�������D���ł���B��w�̌��J�u���̎��ɂ͏����ȊȒP�Ȏ����ł��q���B�̎Q����]�҂��E�����A
�L���L���P���ڂ����邱�Ƃ��ł���B���ݓI�ɂ͍����D���̒j�̎q�͂�������̂ŁA��l�����̂悤�Ȏw�������Ă݂Ă͂ǂ����낤�B��O�̑̌��ŕs�v�c��
�Ɗ������������ɏ������߂Ă�������B�s�v�c�Ȏ����̐����闝�R��J�j�Y���ɂ��ĉ����𗧂Ă�����B���������������Ƃ������ɂ͂ǂ̂悤�Ȏ�����
�����炢�����l��������B���D��┎���ق͎q���B�Ɏ���Ŏ��������A���ɂP�A�Q������̎w�����s�Ȃ��B
�@��
���ȑ��u��ޗ����g�킸�A����
�łł�������͂�������B�ŋ߂��̂��Ƃ����g�Ŏ����ꂽ�͕̂���s���搶�ł���B�u�A�Q�n�`���E�̗c���͕\�ʂ��U���U�������ꏊ��匉��������ɊD�F��
匂ƂȂ�v�Ƃ��������𗧂āA���������Ō����ɏؖ����ꂽ�B������_���I�ɑg���ĂĂ����v���Z�X�����炵���B������ӂɂ���ޗ��������g���ē���ꂽ
�������ʂ��ꗬ�̍��ێG���Ɍf�ڂ��ꂽ�B�������Ƃ��c�ɍs�����ɑ�w�̃L�����p�X����������ŁA�s�v�c�Ɏv�������ɂ������o���킷�B���ꂩ�痧�Ă���
���ɂ́A�Ⴆ�Ύ��̂悤�Ȃ��̂�����B�u�n���r���J�}�L���̗c���͉��F���Ԃ�I��ŏW�܂�v�u�����V���`���E�̖{���̐����n�͍r��n�ł���v�u�S�}�_��
�`���E�̗c��������z�~����͖̂̍����ɗ��t���Ȃ����ł���v�B�������ؖ����邽�߂̎����͏����l����Αg�ݗ��Ă邱�Ƃ��ł���B���w�������l�܂�
�P�l�Ŏ����Ɏ��g�ނ��Ƃ��ł���B���Ƃ͂����ɃG���K���g�Ɋm���ɏؖ����Ă݂��邩���r�̌����ǂ���ł���B
�@��
����PCR���u��DNA�V�[�P���T�[���g�����Ƃ��D�ꂽ�����ł͂Ȃ��B���
�K�v�Ȃ͎̂��R��s
�v�c�Ɏv��������{�����ƁA���ɉ������ؖ����邽�߂̎����̑g���Ă��l�����邱�Ƃł���B�V�����������N�́u�̏W��������ցv�A���ꂪ���ȍD������āA
�n���͖L���ȉȊw�҂���邱�ƂɂȂ�̂ł͂Ȃ��낤���B�B��̊뜜�́A�̏W�ƕ��ނ������Ă�����l�ɂ��ꂪ�{���Ɏw���ł��邩�Ƃ����_�ł���B
|

���@�F�i
���{����w��w�@���w�n�����ȁ@����
|
2011.5.8
�@No.1 �@
�@�@��
�������҂͂Ȃ��������Ă��邩�H ��@�V�@�ȁ@�O
|
�@���̂Ƃ���A���D���̎�҂�����
���Ȃ��Ȃ��Ȃ��Ă����A�Ƃ����Q�������X�ŕ������B���{�̎R�삪�J�����b�V���ŁA���̂��݂��������������Ƃ����낤���A���ȊO�̒ᑭ�H�ȃQ�[���̔�
���Ȃǂ���҂̋��������Ȃǂ����Ȃ��Ȃ����̂�����Ƃ��Ă������悤�B
�@�������A���͂���ȊO�̂����Ƒ�
���Ȍ����́A������䂪���̍����w��majority���A�ÐF���R�Ƃ����L�ڕ��ފw�Ő�
�߂��Ă��邱�Ƃɂ��A�Ǝv���Ă���B�Ⴆ�A�Ԗ�����ߎ���́u����͐ԁv�Ə������A�ʔ������Ƃɂ́A����̎�̐Ԃ͐F�f�n�A�����͍\���F�ł����
�������邵�A�������i�܂��͐ԁj�F�ł��F�f���قȂ�ȂǁA�P�ɋL�ڂ�ǂ����ł͕�����Ȃ��B���̂悤�ȗ�͑��ɂ��R�قǂ���A�ŏI�I�ɂ͂����̐F��
�x�z�����`�q�̓���Ƃ��̔����̐���@�\��m��K�v�����邱�Ƃ͂����܂ł��Ȃ��B���̂悤�Ȏ����ɁA�����̑��l���̈�[���B����Ă���A�i���̋@�\�̉�
���ɂ��v������ł��낤�B���₱���̉�͉͂\�ƂȂ����B
�@���ꂾ�������w�������̐i������
���A���Z�Ȃǂ̋���̏�ł��A���Ȃ荂�x�ȐV���������w�������Ă���̂�����A��قǂ̗�O�������A�����A���݂̓��{�́g�������w�h�̌����@�ւɓ�����
�ꐶ�����������Ƃ����҂��������A����ɔ����A�A�}�`���A�̒����̖ړI�ӎ��ɂ��ω��������͓̂��R�̋A���ł���B�^�S�����̍�N�x�̍b���̉�ڂ̍���
�݂�ƁA��N�L�ڂ��ꂽ�V��̃��X�g����̂ŁA����DNA������Ŏ��X�Ƃ������g�{���ɍ�
���͖ʔ����h�@�Ǝ����v�������͂قƂ�Ǐo�Ă�
�Ȃ��B
�@���́A���ފw�͕K�v�s���Ȃ���
���Ǝv���Ă͂��邪�A�����܂Ői�����������w�S�̂���݂�ƁA����܂ł̕��ފw��10%���炢����A�����w�S�́i������
��ʓI�ɂ����A�����w�j����݂ď\���ł���Ǝv���B����ɂ́A�����ق��w�̍����������̒�������Ȃ��Ƃ����{�ɑ����A���ފw�҂����Ń|�X�g��Ɛ肵��
�����Ƃ��K�v�ł���B��������Β��D���̎�҂̑����́A�����ɂ͂����ƁA�����Ɩʔ������Ƃ��R�قǂ���̂�����A���낢��ȐV�����Z�p��l������g�ɂ�
���A�V���������w�̓��i�݂����Ɗ�����̂ł͂Ȃ����Ǝv���B�����w�����������i60�N���́j�A�̍]��s��v�搶���u��
�吨�̂ЂƂ�����Ă��邱�Ƃ͂��ȁB�����Ŗʔ������Ƃ��݂��A�����ʔ�������v�Ƃ���ꂽ���A���⍩���w�ɂƂ��āA���ꂪ�d�v�Ȃ̂ł���B�v����
�ɁA�l�܂˂͂���ȁB�����œ����J���A�Ƃ������Ƃł���B
�@�����̓r�����Ȃ����l������݂�
�A������Ȃ����Ƃ��炯�ł���B�����͑S������majority���߂�̂�����A���������ł͐i
���̖{����������Ȃ��Ǝv���B���̂Ƃ���A���f���������g���āA�V�������ۂ�A����܂őz�����ł��Ȃ������悤�ȋZ�p�������Əo�Ă��Ă���B�����^
������A�Ƃ����̂ł͂Ȃ��B�Ⴂ����̕��͂��������܂����p���A���ǂ��A�X�ɂ͎��g�œƎ��Ȃ����������āA�ʔ������̐��E�����ۂƂ��Č��邾���ł�
���A�ǂ̂悤�Ȏd�g�݂ł��̂悤�Ȃ��Ƃ��N���Ă���̂��A����������A���ۂ̖{�����𖾂��Ăق����B�����Ȃ��Ďn�߂č����w�͐����Ȋw�����ԂƔF�߂��
��̂ł���B���̂悤�Ȋϓ_����݂āA����DNA������̖����͂��ꂩ��܂��܂���
�傫���Ȃ�ł��낤�B
|

���V�@�ȎO
������w�E�L����w���_����
|
|
2015.02.15
No.16 �����ƐA�� ���J���@����
�@�ǂ������킯���A���̐S�t����
���炸���ƐA�����D���������̂ŁA�����̖��m�ȋL���͏��w�Z��N���̉ċx�݂̎��R�����ł��B�i�{�[�����ɖ�R�̊G��`���A���ꂼ��̍����̐���ꏊ�ɒ���
�z�u���܂����B1970�N�㏉���͐��Ԃ̒��Ő����𗝉�����Ƃ��������������̂ł��̉e���������̂��ȂƎv���܂��B�����ɏZ��ł����̂ŁA�I�I�����T�L��
�ǂ�����N�k�M�̖��������̂ł����A��������̃J�u�g���V��N���K�^�͂��炸�A�J�i�u������B���̂����A�F�B�̉Ƃɂ������H���A���ɐS��D���A����
�Ɏ����Ă��܂��B
�@��w�͐A���w�����ő�w�@�̌������͐A�����ɂ������̂ŁA�w���ō����i��搶�̖��Ғœ����w����u���āAR.M. Alexander����́uThe
Invertebrates�v��ǂ�Ŗ��Ғœ����̑��l���ɖ��f���ꂽ�̂ƁA�ΐ쓝�搶�̃[�~�Ŋ���t�Y�搶�́u�����̐����w�v�N�����ēǂ�Ŗʔ���
�ȂƎv�������炢�ŁA�����Ƃ̐[���������͂���܂���ł����B��w�̃T�[�N���͐����w������ŁA�����������̍�{���炭���c�ӏG�V����A�N���K�^����
�G�ؗ�����A���S�ʂ̐[�Õ��n����Ȃǒ����͂��������̂ł����A�����ɍs���Ă����[�~�X�V�W�~���̓i�K�T�L�V�_���h�L�ɖ��͂������Ă��܂����B
�@���ɋ����������������͉̂������Ċ���ɗ��Ă���ł��B���C���Ă��炭���āA�����̖ї��G�Y�搶�����{�ɗ��āu���J���N�A�`���E���Ȃ����v�Ƃ���
�Ȃ肨��������āA����s�y�b�g�}���������āA�Z�p�x�����̕S�X�R��q����Ɏ����������o���܂����B���̌�A���̎O�}�L���搶����W���u�`�̋@�������
�����A�f���Ă��b�����Ă��邤���ɁA�O�}�搶���Ȃ̑c���̍]���O�搶�̂���q����ł��邱�Ƃ��킩��A�܂��A�O�}�搶���N���}�`�X�̈��ƂŐH���Ƃ���
�ȏ�ɐA���ɐ��ʂ���Ă������Ƃ�����A���̌�A���X���w�����������Ă��܂��B�܂��A��㏟�炭����͂��ߑ����̂���q����ɂ��䐢�b�ɂȂ��Ă��܂��B��
���搶�́A���{�ŕ��s�I�ɐ����Ă����`���E��DNA��p�����n����̓O���[�v�����܂��������ł��������Ȃ����ƍl�����A����DNA�������ݗ���
��܂����B���̉�͒��ނɌ��炸�A�����S�ʂɂ܂�����A�w���w���̂��납�炢�낢�낲�������������Ă������q�i���w�̑��V�ȎO�搶��A���ƂŏW�c��`�w��
����͂�����������{�b��搶�̍����w�҂Ƃ��Ă̈�ʂ��͂��߁A�����̂��Ƃ��w���Ă��������ƂĂ��L�v�ł����B�n�ӈ�Y�搶�ɂ́A�M�t�`���E�̏W�ɘA��
�čs���Ă��������ăJ���A�I�C�̐i���ɋ���������������������Ă�����������A���̌�����낢��Ȏ��Ō䐢�b�ɂȂ�܂����B
�@�����ʂł́A����n�a�̈��`���a�����A���w�����̈��y�̒˒J�T�ꂭ��Ɠ��{�ł͂��߂ẴG�{�f�{�̃V���|�W�E�����u�����ƐA���̔����̈Ⴂ�v��
�����e�[�}�ŊJ�Â��A������A���Ɣ�r���邱�Ƃ̖ʔ����ɋC�Â��܂����B�����āA����̏�쒼�l����\�����ꂽ���������̓���̈�ŁA�n�i�J�}�L��
�ƃR�I���M����������Ă�����n�����搶�̔ǂɉ����Ă��������A�����̕��X�ƂT�N�ԓ��������O���[�v�ɏ������A�����̎h�����܂����B���̔lj�c�̎�
�ɁA��D���̑q�J�����������Ɏt���Ƌ��A�lj�c�̋x�ݎ��Ԃ��ɒ����ɗ�݁A���̗���ŁA�������u��`�v�ɓ����ƐA���̔����̈Ⴂ�ɂ���2002
�N����Q�N�ԃ����[�Βk�������Ă��������܂����B���̓��e�A�Ⴓ�y��A���ł��ʔ����_�������̂Ő}���قŒT���Ă݂ĉ������B���̍��A�ړ�����b����������
�ł����A�ړ��������Ȃ������Ȃ́u�ߏ��ɂ����R�������Ă��v�Ƃ������炵�������ɂ��A�ЂƎR�����āi�R�т͂��ǂ낭�قLj����ŋ����قƂ�ǂ������
���̂ł����߂ł��j�A���T�A���̏W�ō������ɂ����������W�߁A�N���K�^�u�[���������Ă��낢��Ȏ�ނ̃N���K�^����Ă܂����B�����āA�s����w�ɏW��
�u�`�Ɏf�����Ƃ��ɂ��낢��i���ɂ��Ă��f���������Ƃ̂������ΐ�Ǖ�搶�́u�����̒a�� :
��疜��ւ̐i���ƕ����i�����V���j�v�ɏo�Ă��鍩�����V�~����`���E�܂ō̂�܂���A�̌n�����č����̕������邱�Ƃ��ł��A�܂��܂��������N���܂�
���B���̍��A�p�v�A�L���C���N���K�^�̐F�ʑ��^�ɋ����������A���{�̔|�{���̒I�����g���Ĉ�l�Ő��S�C�̌�z�������n�߂܂����B�����A�����o������Ɛ��b
�ɍ���̂ƁA�p�シ��ɂ�ߐe��z�̉e�����Ђǂ��A�U��قnjp�サ���Ƃ���Œ��~���Ă��܂��܂����B���̑����͒�N��ɂ�낤�Ǝv���Ă��܂��B
�@�ї��搶�̌����͂ǂ�ǂ�i�W���A�O�}�搶�A��t�G�K����A���؍F�i������n�߂Ƃ����搶���̂����͂̂��ƃZ�Z���`���E�̌n���ɂ��Ă̘_���\�ł�
�܂����iTanikawa-Dodo et al.
2008�j�B���̂���A����Ń[�u���t�B�b�V���Ő������v���������Ă����^��O���N���q���̂��납��n�i�J�}�L���̌������������������ǁA�ǂ�������
�̂��Ƒ��k�ɗ��܂����B����Ȗʔ������̊w�U���ʌ������ɏo�����為�������ʂ�Ɨ�܂����Ƃ���A�����Ȑ\�����������グ�����̑�����܂����B����ɁA�k
��_�w���ɂ����哇�ꐳ�N���A�w�U���ʌ������Ƃ��ăN���~�z�\�K�̃z�X�g���[�X�]���̎d���������ł�肽���Ƃ����Ȃ�K�˂Ă��āA�������ɉ�X�̃Q�m���Z
�p�Ƒg�ݍ��킹��A�z�X�g���[�X�]���̌�����`�q���P���ł������������̂ŁA�w�U���ʌ������ւ̐\���𗹉������Ƃ���A����܂��̗p����܂����B�A����
�W�Ő\�����Ă����������ɂ����̂ł����A�����ʂ�͍̂�������A�������A�ɂ߂Č��I�����[�h���[�J�[�̓�l���A�����肾�����������ɉ����A��
�܂܂łƈ�������͋C�ňꓯ�A�����ɐ���オ�邱�ƂƂȂ�܂����B�哇�N�́A�S�X���������s���A�ї��搶�ƎO�}�搶�����S�ƂȂ��Đi�߂Ă����^�e�n
�`���E�̌n���̘_�����n���ƃz�X�g���[�X�]���̊ϓ_����ʔ����܂Ƃ߂Ă���܂����iOhshima et al.
2010�j�B�n�i�J�}�L���̋[�ԂƃN���~�z�\�K�̃z�X�g���[�X�]��������`�q�͂����������i�s���ł��̂ŁA���ʂ͂��y���݂ɁB�܂��A�哇�N�ƃz�X�g���[�X
�]���̌����ɂ��Đ܁X���낢��c�_���Ă��������ɁA�u�����K���`���̐i���v�̑�\��ł��邱�ƂɋC�Â��܂����B�����K���`���Ƃ́A��̕ψقł͓K���x
���������Ă��܂����A�������̕ψق��d�Ȃ�ƓK���x��������悤�Ɍ�����`���ł��B�Ⴆ�A�z�X�g���[�X�]���́A�c�����V�����z�X�g��H�ׂ���悤��
�Ȃ�ψقƎ��e���V�����z�X�g�ɎY������Ƃ����Q�̕ω����N�����Ă͂��߂Đi�����܂��B�c���Ǝ��e�̗����̌`�����i������A�������A�����z�X�g�ɓ]����
��悤�ɐi�����邱�Ƃ͋ɂ߂č���Ɏv���܂����A�����ł̓z�X�g���[�X�]���͕p�ɂɋN�����Ă���A�Ȃ�Ƃ��s�v�c�ł��B���ꂪ��̂��������ƂȂ��āA
�T�̍b���̑q�J������A�A�Q�n�̋[�Ԃ̓������F����Ɩx���搶�A�����̋����̐[�Â���A�J�C�R�̃z�X�g���[�X�]���̓��c�������ƐV�w�p�̈�𗧂��グ
�邱�Ƃ��ł��܂���
�ihttps://staff.aist.go.jp/t-fukatsu/SGJHome.html�j�B
�@���āA����Ȃ킯�ŁA���͂����ς�A�����S�Ɍ�����i�߂Ă���̂ł����ihttp//www.nibb.ac.jp/evodevo�Ghttp://www.nibb.ac.jp/plantdic/blog/; http//www.nibb.ac.jp/evodevo/tree/00_index.html�j�A
���������A���ɍ����Ɣ�r����Ȃ����Ă��܂��܂����B�A�������Ȃ���A�����ł͂���Ȃ��Ƃ͂�����̂��ȁA�����̉��ɂ�����̂��Ȃ��A�Ƃ����l����
���܂��B�����́A�]�ʈ��q�ɂ���ĂЂƂ����܂�̑g�D����扻���Ĕ������܂����A�A�����Ԃ��`������Ƃ��Ɏ����悤�Ȏd�g�݂��g���Ă��܂��B�܂��A���
���[�𑝂₵�Ă����ҒŌ`���́A�A���̕����g�D���J��Ԃ��t������ĐL�тĂ���l���Ǝ��Ă��܂��B����A�זE�����Z�v�^�[�̕ω��œ����ꏊ��ς��邱��
�ɂ���ĐV�����튯�A�Ⴆ�Ί{���`�������Ƃ����悤�Ȃ��Ƃ́A�זE�������Ȃ��A���ł͖����̂��Ȃƒ��炭�v���Ă��܂����B
�@�Ƃ��낪�A�N�n�߂ɖʔ������Ƃ�����܂����B�_�[�E�B�����r�[�O�����ōq�C�������납��A�`����[��Caltha dionaefolia�i�H
���A���̃n�G�g���\�E�Ɏ��������E�L���J�j�Ƃ����A�����m���Ă��āA�H���A���ł͂Ȃ����Ƃ͂��łɂ킩���Ă���̂ł����A���̊�Ȍ`���ǂ����Ăł���
�̂��s�v�c�Ɏv���Ă��܂����i�}�P�j�B���܂��܁A�`����[�̃i�o���m���ŃR�P�̍��ۊw��������̂ŁA���łɎ����n�����ɍs���܂����B���̃T���v�����
���ώ@����ƁA���Ԃ�A�t�̈ꕔ���Ȃ����Ė����������̂��ȂƎv���Ă��܂����B�܂��A�ߏ��ɕʂ̎�ނ�Caltha appendiculata��
������ނ������Ă��āi�}�Q�j�A������͂���ɖ�����������ŗt�̏ォ��t���������悤�ɂȂ��Ă���̂��悭�킩��܂����B���ꂼ��ȑO�ɕW�{�Ō�������
�͂������̂ł����A�����n�ł��낢��Ȕ����i�K���̕ψق̂���W�c������ƁA������W�{����������̂Ƃ͂��Ȃ������l�����U������Ă��܂��B��ɂ�
�����ӂ������ԎR�̏�ŁA�ӂ��Ǝv���o�����̂������t�c�o�L�ł����B���̉��|�i��A���i�͗t�悪���\���^�Ȃ̂ł����i�}�R�j�A�Ƃ��ǂ��A�t�̐�[�ɂ�
����t��t�����悤�ȗt���`�����܂��i�}�S�j�B����A�ǂ����A�ꂽ�t�̉����������Ƃ��̂悤�ȕ\���^�ɂȂ��Ȃ����Ǝv���Ă����̂ł����A����
�E�L���J�̗t�̖����ƈꏏ�ɍl����ƁA�A���̐V�K�`���i���ɂ���������̖����ׂĂ݂����Ȃ��A�Ǝv���Ă��܂����B����ɁA�H���A���̃E�c�{�J�Y���̓�
�R�ψّ̂ő܂������ɊJ���Ă��܂����̂��������Ƃ�����A���������Ă���������Ői�������̂��ȂǂȂǁA���낢��Ȃ��Ƃ��v���o����Ă��āA�����킸�A
�ɂ�ɂ₵�Ă��܂��܂����B����ɁA�����̍זE�ړ����āA�v�́A�זE���m�̐ڐG��ς��邱�Ƃ����ǁA�A���̖������č��܂ŐڐG���Ȃ������זE���ڐG���ĐV
�����`���ł��邱�Ƃ�����A���́A�d�g�݂Ƃ��Ă͓������Ƃ����A�ƋC�Â��A�����킸�A���������A�ƌ��ɏo���Ă��܂��܂����B
�@����A�A�����ٓI���Ȃ��Ďv���锭���l�������邩�ȂƎv���܂��B�i�o���m���ł͓d�q���C�����ǂ߂Ȃ������̂ł����A�g�����W�b�g�̎��ɁA���܂��܉^�ǂ�
�k�ĂɎ�������H���A���T���Z�j�A�i�}�T�j�̕ߒ��t�`���̘_���̃��r�W�������ʂ��͂��A��X�������Č������A�Ή���M�����҂̕�����������ɂ��肢���܂�
���B���̘_���iFukushima et al.
accepted�j�́A�T���Z�j�A�̕ߒ��t�͗t����̓���̏ꏊ�ōזE����������ω������邱�Ƃő܂��ł���Ƃ����A�זE�����ʐ��䂪�܂��܂���Ĉ�
�Ȃ����K�w�ł̑傫�Ȍ`�ԕω��������N�������Ƃ����������܂Ƃ߂����̂ł��B�A���זE�͓����Ȃ��̂ōזE�����ʂ̐��䂪��̌`�Ԍ`���ɑ傫�ȉe����^����
���B�����ł��זE�^��������Ă���g�D������̂ŁA���Ԃ�A�����悤�Ȏd�g�݂Ői�������튯�������Ƃ��肻���ł��B�����A���̂悤�ɕ���O�̂��̂��A
�����S�̂̕������������̂͂قƂ�Ǖs�\�ł��B����ȂƂ��́A���m��ɑ��k����̂���ԁB����DNA������́A�ӂ���͐ړ_�̂��܂薳�������W�̐搶
���ɂ���ł��A���낢�닳���Ă��炦��̂ŁA���������̂Ȃ��m�I�D��S�[���̋@��ɂȂ��Ă��܂��B���ЁA����Ƃ���낵�����肢�\���グ�܂��B
Fukushima, K., Fujita, H., Yamaguchi, T., Kawaguchi, M., Tsukaya, H.,
and Hasebe, M. Oriented cell division shapes carnivorous pitcher leaves
of Sarracenia purpurea. Nat. Com. Accepted.
Ohshima, I., Tanikawa-Dodo, Y., Saigusa, T., Nishiyama, T., Kitani, M.,
Hasebe, M., and Mohri, H. (2010). Phylogeny, biogeography, and
host-plant association in the subfamily Apaturinae (Insecta:
Lepidoptera: Nymphalidae) inferred from eight nuclear and seven
mitochondrial genes. Mol. Phylogenet. Evol. 57: 1026-1036.
Tanikawa-Dodo, Y., Saigusa, T., Chiba, H., Nishiyama, T., Hirowatari,
T., Ishii, M., Yagi, T., Hasebe, M., and Mohri, H. (2008). Molecular
phylogeny of Japanese skippers (Lepidoptera, Hesperiidae) based on
mitochondrial ND5 and CO1 gene sequences. Trans. Lepid. Soc. Japan 59:
29-41.
�}�P Caltha dionaefolia
�}�Q�@Caltha appendiculata�̂P���̗t�B
�}�R�@�����t�ւ̐�[���ꂱ�t
�}�S�@�����t�ւ̗t�̐�[�ɂ�����t���ł����悤�ȗt
�}�T�@�T���Z�j�ASarracenia purpurea�̂P���̗t
|
���q�A����Drymis�ɂ����x�I
�`���암�ɂ�
���J���@����
�@
��b�����w�������@�����i���������勳��
|
2014.07.08
No.15 �x�j�V�W�~�ނ̓�
���@����
�@�x
�j�V�W�~�Ƃ���������������
���낤���B������c���̘H�T�Ȃǂł悭�����鏬���Ȟ�F�̃V�W�~�`���E�ł���B���̒��Ԃ͐��E���炨�悻100�킪�m���Ă���A���̑����͑������ł�
�邪�A�M�ѓK���̎�ł͐X�ѐ��̑�������B�c���͎�Ƀ^�f�ȐA����H���Ƃ��邪�A�k���ł͑S���މ��̂Ȃ��A���Ɉˑ�������̂�����B���`�����͂��Ȃ葽
�l�ŁA�ꕔ�ł͋[�Ԃ������A���������̑��l���͗Y����퓙�̌`�Ԃ̕ω��Ƃ͂����Ί֘A�������Ȃ��B�܂��A�x�j�V�W�~�ނ̕��z�͂��Ȃ���قŁA���
���[���V�A�嗤��k�ĂɎY������̂́A�{���l�I�A�p�v�A�j���[�M�j�A�A���āA��A�t���J�암�A�j���[�W�[�����h�̂悤�Ȓn��ɏ����̎킪�u�����z���Ă�
��B�Ƃ��낪���̃O���[�v�̓�͂��ꂾ���ł͂Ȃ��B��L���������`�E����ƗY�����Ƃ̊W�����łȂ��A�����̌`�ԕω��ƒn���I���z�Ƃ̊֘A���������S
���Ȃ����̂���ŁA�n���I�Ȍq���肪�����ς茩���Ă��Ȃ��̂ł���B
�@���̕s�v�c���ɍŏ��ɋC�t�����̂́A�u�\����`�����w�v�Œ����Ȍ́E�ĒJ�čO�搶�ł������B1974�N�ɐ搶�͂���܂Ńx�j�V�W�~�ނ̕��z���S���m���
�Ă��Ȃ������p�v�A�j���[�M�j�A����V��Melanolycaena��
�n�݂���2�V��M. altimontana��M. thecloides��
�L�ڂ��A���킹�Đ��E�̎�ȃx�j�V�W�~�ނɊւ��鍂�����ނ̍Č������s�����B���̐܂ɖ{�Q����`�Ԃ�ԁA�n���I���z�Ƃ̊֘A�ł��̂悤�Ȕ�߂������
���o���A�𖾂ł���u�i���v��u�K���v�A�u�[�ԁv�Ƃ�������ʓI���������̂ɎQ�l�ɂȂ�ƍl�����i�ĒJ,
1999�j�B���̖������炯�̃O���[�v�ɂ����Ƌ��������������Ă���ꂽ�悤�����A���̓�������������Ȃ��܂܂ł������B
�@1998�N�ɏC�m�@���Ƃ��ċ�B��w�̑�w�@�i�w�������́A�����̐��b�l�w�������ł����������̌`�Ԋw����ьn���w�̌��ЁA�O�}�L���搶����̊���
�ŁA���̃x�j�V�W�~�ނ̍������ނ̍Č����ƌn���i���w�I�Ȍ������C�m�_���̃e�[�}��I�B�������A�O�}�搶���ĒJ�搶���炱�̕s���ȓ���Ă���
�̂����������ł���B���̌������s���ɂ�����A�܂��͎ĒJ�搶����̏��������炤���ƁA���킹�Ė{�����ւ̋��͂�v�����邱�Ƃ��O�}�搶���炲�w������
���B�����A�ĒJ�搶�Ɏ莆�������Ă����̂��肢��Ԃ����莆�𑗕t�����Ƃ���A�Ԃ��Ȃ����J�Ȃ��Ԏ����͂����B�����ɂ͎�����̂��肢�Ɋւ�������ƁA
�����g�̃R���N�V��������������Ă��镺�Ɍ����l�Ǝ��R�̔����قɑ����^��ŕW�{�������������Ƃ�������Ă������B���̌�A����ɓ��̎莆�Ɠd�b�̂�
����𑱂��A�ĒJ�搶�ƕ��Ɍ����Œ��ڂ�����Ė{�����̖��ƓW�]���������������B�ĒJ�搶�ƌ����A�x�j�V�W�~�ނ����邱�ƂȂ���[�t�B���X�̌���
�ł���������z���A�t�W�~�h���V�W�~�̑����ɂ���������Ă���`���E�ތ����̑�Ƃł���B��������ۏo���̃`���E�����������ɂƂ��āA�ĒJ�搶�͍\����`
�����w�����������_�w�҂Ƃ��Ă����A�ނ���`���E�ނ̕��ފw�҂Ƃ��Ă�������ْ��Ɗ������̕��������������Ƃ������Ă���B
�@��N����A���͏C�m�_�����Ȃ�Ƃ��܂Ƃߏグ�A�x�j�V�W�~�ނ̌n�����ފw�I�����ɂ��Ă͈ꉞ�̋�肪�������A���͌n���̕����ł͂������肱�Ȃ���
�a�����������B�܂�A�`�ԃf�[�^��p�����n����̓\�t�g�ɂ��Őߖ�@�œ����ꂽ���ʂɂ��܂莩�M���Ȃ������̂ł���B����ł��A�ĒJ�搶�ɂ͂�������
�Ă��ӌ����f�������Ǝv���A���҂ƕs�������荬�������C�����ŏC�_��o�̑O�ɂ��̓��e��X�������B�Ԃ��Ȃ����āA�ĒJ�搶�����ʂ̎莆���͂����B������
�́u�E�E�E���̂����p�l�ɂȂ�̂ł͂Ȃ����ƁA�����ꂨ�̂̂��Ă��܂��B�E�E�E���������\�͂�����������܂��B�������݂܂���A����ȏ�̓����̂悤
�ł��B�v�ƋL����Ă������B����܂Ŏ莆�̂�����搶�Ƒ����Ă������ŁA�������ɂ��̂悤�ȕ��͂͌������Ƃ��Ȃ��A�����ɐ搶�̑̒���S�z����C����
�����ݏグ���B���̍��ɂ́A���łɋC�͂�̗͂������Ă����̂ł��낤�B���̈���ŁA�搶�̂����҂ɂ������o���Ȃ������̂�������Ȃ��Ƃ������]�����P��
���B�搶�̂��l���ȏ�̂��̂�E���Ă��Ȃ������̂ł��낤���B
�@���̌�A���m�ے��ɐi���́A�e�[�}���V�W�~�`���E�ȑS�̗̂c���`�Ԃƌn���w�Ɋւ��錤���ɍL�������߁A�x�j�V�W�~�ނ̌���������ɒNj����邱�Ƃ͂�
�������B����ł��n���W�̈�a����s�����͂����Ɛ@���Ȃ��������߁A�@�����ΐ���Ƃ����q�n���w�I�ȃA�v���[�`����`�Ԃœ���ꂽ���ʂ���������
�ƍl���Ă����B�w�ʎ擾��A���͊�b�����w�������̒��J�����א搶�̏Љ�ŁA���Y�L�ނ̕��q�n���w�I�����Œm���铌����w�̏㓇��搶�̂Ƃ���Ƀ|�X�h
�N�Ōٗp���ꂽ�B�Ζ����ԊO�͎��g�̌��������Ă��ǂ������̂ŁA����̓`�����X�Ƒ����x�j�V�W�~�ނ̕��q�n����͂��n�߂邱�Ƃɂ����B���̎��̂��߂�
DNA�T���v�������͗��ߑ����Ă����̂ł���B�~�g�R���h���A�Ɗj�̑o����DNA����n����͂�i�߂Ȃ���A�s�����Ă���T���v�������߂ăA�t���J�⒆��
�O�@�e�}���Ȃǂɂ��K��A�悤�₭�x�j�V�W�~�ނ̍����n���ɂ�����S�̑����������̂́A�{�����̒��肩���10�N��̂��Ƃł������B
�@���ʂ��o�Ă݂�A�n���n���w�I�Ȋϓ_����ł����[�Y�i�u���ɉ��߂ł�����̂ł������B�Ⴆ�A�p�v�A�j���[�M�j�A�ɐ�������Melanolycaena�������̔����O�r跗�߂̌`�Ԃ��璆�Ă�Iophanus���ɋ߉���������Ȃ��Ƃ���Ă������A���q�n���ł͂ނ���O
�ς��قȂ�j���[�W�[�����h�̃O���[�v�Ǝo���Q���`�����A����ɑo�������킹����Q�̓`�x�b�g�⒆�������̍��n�ɐ�������Helleia��
�ɋ߉��Ƃ����i�W���iprogression
rule�j�łقڐ����ł���B�ł́A���q�n����͂ɂ�錋�ʂ����ۂɐ��������ǂ��������ƂȂ邪�A������Č`�Ԃ��������ƁA�s�v�c�Ȃ��Ƃɍ��܂ŋC
�t���Ȃ��������L�h���`�������X�ƌ����n�߂��B���̂悤�ɕ��q�n���̐�������������Ɠ����ɁA�`�Ԃ̎��ʂ╽�s�i���͑z�����ĒZ���ԂŋN���邱�Ƃ���
����A��d�̋����������オ�����̂ł���@�B
�@�ĒJ�搶�̐��O�ɂ��̌��ʂ������������������̂����A���ꂪ����Ȃ��̂��B��̐S�c��ƂȂ����B�܂��C�̂��̂Ƃ��R�̂��̂Ƃ��o���Ȃ����̏C�m�@��
���������ɁA�����̂��Ƃ��������������������łȂ��A����܂ŏ������߂Ă����V�W�~�`���E�Ȃ̗Y�����̌��}�܂ł�搶�͎��ɑ����ꂽ�B�ŋ߁A�����̕`
�}�߂ẮA�{����鎑�i�����邾���̌����҂ɐ���Ă��邩�ǂ������������g�ɖ₤�ƁA�܂������������Ă��Ȃ����ƂɋC�t���A�Ȃ���w�̓w�͂��K�v
�ł��邱�Ƃ�F������̂ł���B
|
���@����
�@
������w�������������ف@����
|
2014.02.07
No.14 �@���A�ɓ��������ʔ���
�r�J�@�M�Y
�@
�������߂ĕ��ƈɓ������̔��䓇�ɍ̏W�ɖK�ꂽ�̂́A������30�N�ȏ�O�̒��w�R�N�̉ċx�݂������B�����A�J�~�L�����V�ɑ傫�ȋ���������Ă������ɂƂ�
�āA�n�`�W���E�g�Q�E�X�o�J�~�L����n�`�W���E�R�o�l�J�~���ȂǑ����̔��䓇�ŗL�̃J�~�L�����V�̒��i�Ƃ̏o��͂܂��ɋ���̘A���������B�������A����
�����Ă��̍K���ȏ͈�ς���B�@
�u���̓��ɂ̓m�R�M���N���K�^����R�����B�v�h�̂���l���炻�̈ꌾ�����̂͏����̗[�H�̎��������B���̎��ɂ́A�{�y���炱��Ȃɗ���
�����ɂ��m�R�M���N���K�^������̂��Ə��X�����͂������A�V�����{�Ƃ����N���K�^���V�̏W�̃x�X�g�V�[�Y���Ǝv��ꂽ�����ł����������ɁA��R����̂ł�
��Α؍ݒ��ɊȒP�ɍ̏W�ł�����̂ƁA�����A���������Ă����B�������Ȃ���A��̓��Ό����ł��m�R�M���N���K�^�͑S���p�������Ȃ��B�R���ڂ̒��Ԃɓ���
�ōł��ǍD�ȐX�ъ����c���Ă���O���R�̒������т��ѓ���ŁA�m�R�M���N���K�^�Ǝv�������̈ꕔ���E�����B�����Ȃ�Ɛ�����ł��������m�R�M���N���K
�^�̎p���������Ȃ����B�Ō�̔ӂ͖ڕW���m�R�M���N���K�^��{�ɍi���ĕK���̓��ΒT�������s�����i���͒����ł��Ȃ��̂ɁA���̂��߂ɖ�ʂ��Ń����^�J�[��
�^�]����H�ڂɂȂ����j�ɂ��ւ�炸�A���ʂ͎S�s�ɏI�����B�@���������̒��A�ǂ����Ă����߂���Ȃ����́u�Ō�̓q���v�Ə̂��āA�a�镃��������A�h����
�킴�킴���������Č��̎O���R�̗ѓ����ċ�`�Ɍ������Ă�������B�������A�肢���ނȂ����O���Ƀm�R�M���N���K�^�����̈ꕔ���E�����ꏊ���߂��Ă�
�����ڗ��������ʂ͂Ȃ��B��������A��`�̂����ꋽ�̏W����������ƕ��͎Ԃ̃X�s�[�h���グ���B�ƁA���̎��A�����^�J�[�̎ԑ�����H��ɂЂ�����Ԃ���
�����N���K�^���V�̎p���͂�����ƌ������B�u�����I�v�������Ԃ��������A���͎Ԃ���~������������h�A���J���ĊO�ɔ�яo�����B�Ō�̍Ō�ɂ���Ƃ̎v
���Ŏ��ɂ������m�R�M���N���K�^�̏��^�̗Y���E�����Ƃ��ł����u�Ԃ������B���̎��̋����͍��ł��͂�����Ɗo���Ă���B�������P���̂��������ɂ��������^
�̗Y�ł͂��������A���܂�ď��߂Č������䓇�̃m�R�M���N���K�^�͐^�����ŁA�܂��Ɉٍ�������Ղ�A����e����ł����{�y�̃m�R�M���N���K�^�Ƃ͑S����
�Ȃ������݂Ɍ������B
�@���̌�A�w�Z�̐}���قɂ������k���فi���a38�N���s�j�́u���F���{�����嚤�ّ�Q���i�b���ҁj�v�ׂĂ݂�ƁA���̔��䓇�̃m�R�M���N
���K�^�͍̏W���̒����ǂ���A�n�`�W���E�m�R�M���N���K�^ssp.
hachijioensis�i�����̎��ɂ͂��̕\�L�̈Ӗ����番����Ȃ������j�ƌĂ�锪�䓇�ƎO��ɂ����Ȃ��ɂ߂ē��ʂȂ��̂ł��邱�ƁA���̈��
�Ŗ{�y�̃m�R�M���Ƃ̐��Ԃ̈Ⴂ�ȂǂɊւ��Ă͑S���𖾂���Ă��Ȃ����Ƃ�m�����B�u���������̓��ʂȃm�R�M���N���K�^���v�������ɍ̏W���āA���̐��Ԃ�
�𖾂������I�v����ȗ��A���̔��䓇�w���n�܂����B
�@�K�����̎��Ɓi���m���j�̗��e�͕��C��`�Łi����������ƕ��͐�̔��䓇�ł̌o���Œ��肽�̂�������Ȃ����j�A�����N�̎����P�ƂŁA������
�w�����Ɂi�傫�Ȑ��ł͌����Ȃ������R�w�Z���x��Łj�����̍̏W�n�ɒʂ����Ƃ��ٔF���Ă��ꂽ�B���A�Œ��w�E���Z����̖K����ʂ��āA�����̃n�`�W���E�m
�R�M���N���K�^���̏W�ł��������łȂ��A�����̔��������͖{�y�Y���������Ƒ����T�`�U���ł���{�y�ł̌o������x�X�g�V�[�Y���Ǝv���Ă����V�����{�ł�
�x�߂������ƁA���s�X����������Ԃɂ��������邪���ɂ͏W�܂�Ȃ����ƁA�c���͌͂ꊔ�̍�����|�̐ڒn�ʂɐ��E���邱�ƁA�Y���|�̉��œ꒣���
�Y���ɂ��鎓�ƌ�����邽�߂ɑ҂��\���Ă��邱�ƁA�Ȃǃn�`�W���E�m�R�M���N���K�^�Ɋւ��鐔�����̐V���Ȓm���邱�Ƃ��ł����B�n�`�W���E�m�R�M���N
���K�^�ȊO�ɂ��A�n�`�W���E�l�u�g�N���K�^�ⓖ���܂��L�^�̂قƂ�ǂȂ������q���^�N���K�^��R�N���K�^���̏W���A���Љ�i�Ƒ������j�𑗂�`�r�N���K
�^�̋����[�����Ԃ��𖾂��邱�Ƃ��ł����B��w���ɂȂ��Ă���͔��䓇�����łȂ��A�䑠����O��ɂ��n��A�~�N���~���}�N���K�^��}���N���K�^�̍̏W��
���������B
�@���Ȃ݂ɁA���ɂƂ��Ă̏��߂Ă̕ł���u���䓇�ɂ�����L�{�V�J�~�L���̍̏W��v�Ƃ����Z��i�����ނ� No.
190:21-22.�j�́A���̍��ɔ��䓇�œ���ꂽ�W�{�Ɋ�Â����̂ł��������A�n�`�W���E�m�R�M���N���K�^��`�r�N���K�^�̐��Ԋώ@�̌��ʂ́A���̌�
�̏C�m�_���u�N���K�^���V�ȍb���̔ɐB���ԁv�̎�v�����ƂȂ����B�������Č���ƒ��w�`��w���{�ے��܂ł̍ł������Ȏ����Ɍo���������̈�A�̈ɓ������w
�́A�܂��ɂ��̌�̍��������҂Ƃ��Ă̎��̌��_�ƂȂ����ƌ����Ă��ߌ��ł͂Ȃ��B
�@80�N��㔼����90�N�㔼�ɂ����āA������N���K�^���V�u�[����w�i�ɁA�ɓ������̃N���K�^���V�Ɋւ��镪�ފw�I�Ȍ������}���ɐi�݁A�n�`�W���E
�q���^�N���K�^��n�`�W���E�R�N���K�^�A�C�Y�~���}�N���K�^�Ȃǂ̐V�^�N�T�����X�ɋL�ڂ��ꂽ�B�n�`�W���E�m�R�M���N���K�^�Ɋւ��Ă��A���̓Ǝ��̌`�Ԃ�
���ԓI��������u�䑠���Ȗk�̖{�y�n�m�R�M���N���K�^�Ƃ͕ʌn���̔��䓇�ŗL�̓Ɨ���v�Ƃ��鋻���[�������������ꂽ�B����A���̍��̎��͐�痮����
�}���[�V�A���͂��߂Ƃ���C�O�̒����ɖv�����A�ɓ������Ƃ͂�������a���ɂȂ��Ă����B
�@�������A���N�O�A�v��ʓW�J���玄�̒��̈ɓ������ւ̏�M���ĔR���邱�ƂƂȂ����B���������̓~�g�R���h���ADNA��p���������̃m�R
�M���N���K�^�ނɊւ���n���n���w�I��͂������B��͂ɍۂ��āA���B�̈�����܂ޖ{�y�e�n�̃m�R�M���N���K�^�ƃn�`�W���E�m�R�M���N���K�^�����������A
���ʂ�O�A���́A�n�`�W���E�m�R�M���N���K�^���{�y�̃m�R�M���N���K�^�Ƃ͕ʌn���̎�ł���A����Έ⑶�ŗL��Ƃ��Ĕ��䓇�Ɋu�����z�������̂ł���
���Ƃ����q�ł����t������Ɨ\�z���Ă����B�n�`�W���E�m�R�M���N���K�^�̎o���Q���Ƃ��āA���̌`�Ԃ̗ގ������璆�����p�ɕ��z����}���o�l�m�R�M��
�N���K�^��z�肵�Ă������ł���B���������ʂ͗\�z�O�̂��̂������B�n�`�W���E�m�R�M���N���K�^�͈ɓ������̈ꕔ�̃m�R�M���N���K�^�̌Q�ƒP�n���Q����
���A���������̕���̒��x�͂��̑��̖{�y�Y�m�R�M���N���K�^�̒n��̌Q�Ԃ̕ψقƑ卷�Ȃ��������̂������̂��B�����Ď��{�����n�`�W���E�m�R�M���N
���K�^�Ɩ{�y�Y�m�R�M���N���K�^�̏ڍׂȌ`�ԑ���w�I��͂̌��ʂ�����A�n�`�W���E�m�R�M���N���K�^�ɂ݂���`�Ԃ�Ԗʂł̓��ꉻ�́A���t�Ƃ������
�I�ȉa���ԂŌ��܂�����K�v���Ȃ��Ȃ������߂ɁA�R�X�g�̂������A�S�̐L���������߂Ă��܂������ʂƂ��ċ}���ɐ��������̂ł��낤���Ƃ���������
���i�����̉�͌��ʂ̏ڍׂ�2009�N���s�̍���DNA�j���[�X���^�[NO. 2�ŏЉ�Ă���j�B
�@�܂����A����܂ł����͂��Ȍ̂����̏W����Ă��炸�A�قƂ�ǒm���̓����Ă��Ȃ������O���䑠���̃m�R�M���N���K�^�i�����̏W����
���炸�A���R�A��L�̕��q�n����͂ɂ��g�p�ł��Ă��Ȃ��j����������A���ꂼ�ꂪ�{�y�̌̌Q�Ƃ͕ʈ���Ƃ��ċL�ڂ����Ƃ��������i�H�j�����������B��
���Ȃ�ƎO���䑠���̌̌Q���܂ވɓ������S��̃m�R�M���N���K�^�̕��q�n���n���w�I��͂����{�����ɂ͂����Ȃ����A�~���}�N���K�^��q���^�N���K
�^�A�R�N���K�^�Ȃǂ̉�͌��ʂ��傢�ɋC�ɂȂ��Ă����B�K���A�������o�傹�˂Ȃ�Ȃ������̂Ƃ͈���āA���̓w���R�v�^�[�H������������ĒZ������ň�
�������e���ɏo��������B���ꂩ��͌����̍��Ԃ�D���āA�������ɂ͌��n�؍ݐ����ԂƂ������d�����ŁA�e���̒����Ɖ�̓T���v���̍̏W���J��Ԃ����Ƃ�
�Ȃ����B
�@�������ē���ꂽ�T���v���Ɋ�Â��ɓ������̃N���K�^���V�Ɋւ��镪�q�n���w�I��͂́A���݁A�܂��i�s���̒i�K�ɂ��邪�A�\��I�ȉ�͂���
�͗\�z�O�̋����[�����ʂ������������Ă���B���ɁA�䑠���Ɛ_�Ó��ɂ����������Ă��炸���E�I�Ɍ��Ă����M�����ɓ������̌ŗL��Ƃ��ėL���ȃ~�N���~
���}�N���K�^�̉�͌��ʂɂ̓n�`�W���E�m�R�M�����z����������������B
�@�~�N���~���}�N���K�^�͏��^�̃~���}�N���K�^�œ����̒��ł͑c��I�Ȏ�Ƃ݂Ȃ���Ă���B�`�ԓI�ȓ������璆���암�̉��͗���ɕ��z���郉
�G�g�D�X�~���}�N���K�^��p���[�~���}�N���K�^�Ɠ�����Q�Ɋ܂܂��Ƃ���A�]���̍l�@�ł́A�~�N���~���}�N���K�^�́A�c��킪�����嗤������{�A������
�ɓ������ւƐN��������A���{�{�y�ł͉����̗��R�Ŗł�ł��܂��A�ɓ������̌䑠���Ɛ_�Ó��݂̂ɐ����c�����A������T�^�I�Ȉ⑶�ŗL��ƌ��Ȃ���Ă�
���B���ۂ�DNA��͂̌��ʂ�����{�킪����璆���Y�̎�ɋ߉��ł��邱�Ƃ����t����ꂽ���A���̈���ŁA���ɋ����[�����Ƃɒ����Y�̋߉���ƃ~�N���~
���}�N���K�^�̈�`�I�ȋ����͋����قNj߂��A�~�N���~���}�N���K�^�̑c���̈ɓ������ւ̐N���i���z�g��j�͂����ŋ߂ɐ������\�����������ꂽ�B��X��
���̌��ʂ���A�~�N���~���}�N���K�^�̑c���͈ɓ������Ɋu�����ꂽ�̂ł͂Ȃ��A�É��͂̑�×��ɂ���Ĕ�r�I�V��������ɒ����嗤����ɓ������ɕY����
���\�����������ɂ�ŁA���݁A���̗��t���̂��߂̌������݂Ă���B
�@�����āA�ɓ������ɂ́A�n���n���݂̂Ȃ炸�A�i�������w�I�ɂ����ɋ����[����ނ��L�x�ɂ��邱�Ƃ����炩�ɂȂ��Ă����B�Ⴆ�A�����̉�
���ňɓ������̃N���K�^���V�ł͌ŗL�^�N�T�₻�̋߉����ތQ�̍\�����D��킪�����ɈقȂ�A���������ꂼ��̌ŗL�^�N�T�̐��Ԃ���j�����Ԃő傫����
�Ȃ��Ă���Ȃǔ��ɋ����[�����ۂ��ώ@����邱�Ƃ����������B�����������Ԃ̌Q�W�\���̍��ق�e���̌̌Q�ɌŗL�ȓK���`���̐i���ɂ́A���R�I���͂���
���A�Y�����͂��ߋ����I�ȕ��U�ɂ���ĐN�������c��̌Q�ɓ������n�n�Ҍ��ʂ��`�I�����A����ɂ͈�`�q�������Ԃ̋����ȂǗl�X�ȗv�������G�ɗ���
�����Ă���炵�����Ƃ����肳�ꂽ�B�܂��A�Â�����c�_�̂������Èɓ��������ɑ����āA�ŋ߂ł̓t�B���s���C�v���[�g�̖k��ɔ����ɓ������̊�̖̂{�B
�ւ̏Փ˂ƒO��R�n�̌`���ȂLjɓ������̐����n���w�ɖ��ڂɊW����n���Ȋw�I�Ȓm�����~�ς���Ă������Ƃ�����̈ɓ������̐������Ɋւ��錤����傫��
�i�W������v���ƂȂ邾�낤�B
�@���m��Ƌ��k��̋��E�Ɉʒu���A���m�̃K���p�S�X�ƌĂ��قǓƎ��̐����������ł���������A�ŗL��`�����̕�ɂƂ��Ė��������}
�������̏ꍇ�قǂ̔h�肳���Ȃ����߂��A�����Ɏ����ŗL�^�N�T�̕�ɂł���ɂ�������炸�A�ɓ������̍������Ɋւ���n���E�����n���w�I�Ȍ����͑傫
�������x��Ă��銴���ۂ߂Ȃ��B����ŁA�嗤�����f����Č`�����ꂽ�嗤���ł��问���̗l�X�Ȑ����Ɋւ���ߔN�̕��q�n���n���w�I��͂ɂ���āA����
����̓��̐������̌`���ߒ��͓��̐����̎����⏇�ԂƋɂ߂Ă悭��v���邱�Ƃ����炩�Ƃ���Ă���B�������A��������������������Ȃ����A����͂���Ӗ�
�\�z���꓾�铖�R�̌��ʂł���A���f�ƕ��U�i�����I�Ȋg�U�j�̑o�������ݍ����ɓ������̐������̐����̉𖾂͈�ؓ�ł͂����Ȃ��䂦�ɗy���ɖʔ����B����
�����嗤�����f����Č`�����ꂽ�嗤���ł��问�����ΎR���̊C�m���ł���K���p�S�X�ɂȂ��炦�鎖���̂��i���Z���X�ł���A���̓_�ł͂ނ���ɓ�����
�̕�����قǁu�i���̎�����v���铌�m�̃K���p�S�X�ƌĂ��ɂӂ��킵���悤�Ɏv���B
�@���̂悤�ɐ����w�I�ɔ��ɋ����[���ɓ����������A�ɂ߂ėR�X�������ƂɁA�ߔN�A�J���ɂ�鐶�����̔j��ɉ����đ����̊O���킪�N�����A
���̋M�d�ȍݗ����Ԍn�͉�ł̊�@�ɕm���Ă���B���s�X���̋����ɓ������̍����ނɂƂ��Ă̓C�^�`�ƃm�l�R�Ȃǂ̑�^�ٓ��ނɂ��ߐH�͉�œI�ȑŌ���^
������B�C�^�`��m�l�R�ȊO�ɂ��A�V�J��L�����A���M�̐H�Q�ɂ��X�ѐA���̔j����������B�O������Ɋւ��ẮA�����⏬�}���ł͂��̑�X�I
�ɍu�����Ă��邪�A�ɓ���������x��ɂȂ�O�ɁA�ꍏ���������炩�̑���Ƃ�K�v������B���̂��߂ɂ��A�{�e�����������ƂȂ��āA�M�d�Ȉɓ������̐�
�����Ɉ�l�ł������̕��������������A���̑��l���̉𖾂�ۑS�ɌW���Ă��������悤�ɂȂ邱�Ƃ�Ɋ肤����ł���B
|
�r�J�@�M�Y
�@
��B��w��w�@�@��r�Љ�������@�@����
|
2013.06.27
No.13 �@�`���E���̎���
�ї��@�G�Y
�@�O�ɂ����������Ƃ����邪�A���͎q���̍������
�u�`���E���v��
�͂Ȃ��B���ł�
�����ł͂Ȃ��Ǝv���Ă���B�������ɏ����������琶�����D���ł���A�ĂƂ��Ȃ�ΐ^�����ɂȂ��ăZ�~��g���{��L���M���X��ǂ������Ă������A�`���E��
�̂����莔�����肵���L���͂Ȃ��B�t�@�[�u�����Ă��Ă��A�����Ă݂����Ǝv�����̂̓Z�~��A���ł������B
�@���܂ꂽ���N�ɖ��B���ς��N����A����ł��R�����N�Ƃ��Ĉ������ɁA���e���E�ƌR�l�ł������̂ŁA���j�ł��鎄�͉��̋^�����Ȃ����̌�������߂ɗ�
�R�c�N�w�Z�ɓ������B���R���Z��{�����邽�߂̊w�Z�ŁA���w��N����N���I���ē��Z���A�O�N��ɗ\�Ȏm���w�Z�A����Ɏm���w�Z�i���̖h�q��w�Z�ɑ����j��
�i�ނ̂ł���B���������Z��N�ڂɔs��ƂȂ�A������̖ړI�͍�����v���K�^�ɂ��ʂ������ɏI������B
�@���͈�]���čD���Ȃ��Ƃ���낤�Ƃ������ƂɂȂ�A�������Z�ő�w�̗��w���ɓ����w�ȂƂ������̂����邱�Ƃ��͂��߂Ēm���āA����̓��w�Ȃɐi�w��
���B���Ƃ��čŏ��ɏA�E�����̂��O��̗ՊC�������ŁA�����ŃE�j�̐��q�Ŏ������n�߁A�Ȍ㐸�q���߂��鏔���Ɏ��g�ނ��ƂɂȂ����B���̊Ԑ��q�̕ږт�
��v�ȍ\���v�f�ł�������ǂ̎听����V�����^���p�N���Ɠ��肵�ă`���[�u�����Ɩ�������K�^�ɂ��b�܂�A���q�w�Ƃ����w�╪�����邱�Ƃ��ł����B����
�ƍ����͑��̐l�����̌����e�[�}���Ǝv���Ă����B
�@���ă`���E�Ƃ̏o��ł��邪�A���傤�Ǒ�w�����̂���ɉƓ��̐e�����쌧�̐������ɎR�����������B�s���Ă݂�Ƃ��ĕ��R�C���Y���́u�����}�Ӂv��
�����������Ƃ̂Ȃ������`���E��g���{�����������B�����Ŏ���̂��̂����ł��S���̂��Ă݂������̂ƁA�l�\�̎�K���ō����̏W������悤�ɂȂ����B�c
�O�Ȃ���̂�Ȃ��������A�����ɂ̓I�I�E���M���q���E�������܂������悤�ł���B���̌�I�[�X�g�����A�ɗ��w�����ۂɁA�����V�h�j�[�ɂ���ꂽ�ĒJ�čO��
��ɂ��낢��Ǝ�قǂ����A�`���E�ɂ̓|�C���g�Ƃ������̂����邱�Ƃ��w�B���̉��Ŕ��{�b�s����ⓡ���m�v����Ƃ��m�荇���悤�ɂȂ����B
�@������N�ő��Ƃ��ĕ�����w�ɍs������A�����ɂ̓`���E�̗ؕ��𑖍��d���Ŋώ@���Ă����u�`���E���v�̐V��@�ׂ������B�����Ń`���E�̐��q�̓d��
����肩�������A�Ԃ��Ȃ���b�����w�������̏����Ƃ��ĉ���ɕ��C���邱�ƂɂȂ�B��C�ɂ͑�w�̓������Ń~�g�R���h���A�̌��Ђł��钆��@���N�������B
���傤�Ǒ��V�ȎO�������������قŃ~�g�R���h���ADNA�ɂ��I�T���V�̕��q�n�����n�߂�����ł���B
�@�V�삳��͒��V�N�̒�q�̖q�c�T���N�Ƒ��V����Ƀe�N�j�b�N��������āA�M�t�`���E�̋N���ɂ��ă~�g�R���h���ADNA�ň�̓������o�����ƂɂȂ�B
���͊���̏����Ƃ��Ă����ς猤�����̉^�c�ɓw�͂��Ă������A�A���̕��q�n���̃p�C�I�j�A�ł��钷�J�����N������ɕ��C���Ă����@����Ƃ炦�A���V��
�����{����A���؍F�i�����Ƃ������āA�����܂��܂������������������̃V�[�P���T�[���������A�`���E�̕��q�n���̑S���I�ȋ��������𗧂��グ�邱
�Ƃɂ����B
�@���Ă��ꂩ�炪�{��ł���B���̃v���W�F�N�g�ɂ͑S���́u�`���E���v���������͂��Ă��ꂽ�B�����\���N�O�̂��ƂɂȂ�̂ō����͂܂��܂�������
���A���łɑ����̃`���E�����̓V�R�L�O���ł�������A��Ŋ뜜��ł������肷��̂œ��{�Y�̂��̂��W�߂�̂����J�ł������B���Ɉꎞ�͂��ꂪ�Ō�̕W�{
�ɂȂ邩���m��Ȃ��Ǝv��ꂽ�I�K�T�����V�W�~�̍̏W�ɂ��Ă͊��ӂ�����Ȃ��B����ő����́u�`���E���v�̕��X�ƃR�l���������Ƃ��ł����B�������̎���
��������������Ȃ̍̏W�����Ƃ�̂ɑ傢�ɋ��͂��Ă���A�S���犴�ӂ��Ă���B
�@������N�őނ��ĊԂ��Ȃ��A�O�}�L������̎����Ŗ�㏟�炳���ƃS�}�V�W�~�̕��q�n������邱�ƂɂȂ����B����܂ł͂ނ��������̏W�́u�`���E
���v�̊F����ɂ��C�����Ă����̂ł��邪�A�����ŏ��߂Ď��炠�������̏W�ɕ������ƂɂȂ����B���̍ŏ��͗����͂��߂Ƃ���Ìy�����ł������B�����ŃS
�}�V�W�~���̏W�������Ă���k�C���̎R�{��������Əo��A�v���Ƃ͂���Ȃ��̂��Ƃ��̌����ȍ̏W�Ԃ�ɂقƂقƊ��S�����B�`���E�̍̏W�Ɋւ��ẮA����
�����Ă݂āA��コ���R�`�̉i���ÔV�����̗͗ʂɂ��傢�Ɋ����������B
�@�S�}�V�W�~�͖k�C����X����������_�X�Ƃ��Ă������������A�������W�߂�̂���ςł������B���̏\�N�̊Ԃɂ��̂�Ȃ��Ȃ����ꏊ������B�܊p����ɂ�
���̂Ɉ��m���A�É����ł͂��̃v���W�F�N�g���n�܂�O�ɐ�ł��Ă��܂����̂����Ƃ����ɂ����B�����W�{����ł͂܂Ƃ���DNA�����o�ł��Ȃ�����ł���B
����ł��ǂ����嗤�̎������܂߂����͌��ʂ��߂������ɘ_���ɂȂ�Ɗ��҂����悤�ɂȂ����B
�@����Ƃ͕ʂɁA�ߔN�͎������I�ɂ܂��o����Ă��Ȃ��`���E�����߂Ă����炱����ɏo�����Ă���B�����̃`���E�̂قƂ�ǂ��䂪���̐�Ŋ뜜���n���
���ł���B�����ɂ����Ĕ��������u�`���E���v�̊F����̎��͂̂قǂɂ͂܂��ƂɊ��Q���ւ����Ȃ��B�s���|�C���g�ŏꏊ�⎞���������Ă��������̂ł���
���A�����̏ꍇ�������Ă���B�����̂͂��̏ꏊ�ɂ͕K���ƌ����Ă悢�قǕʂ́u�`���E���v�����邱�Ƃł���B��������V�����̃N���}�c�}�L�`���E��
���߂Ď�����߂��̃|�C���g�ɍs�������A�ɓ����v����ɑ������������łȂ��A�����ɓ�A�O���ʂ��Ă���Ƃ��������▼�É��̐l�A�����Ƃ���ꏊ�ő҂��\��
�Ă����҂Ȃǂ������B
�@��コ��̗U���Œ��ފw��Ɋ���o���Ă݂Ă��A���ނ�ԁA���̑��`���E�̂������Ɋւ���u�`���E���v�����̒m���A�����ɂ́A���X�Ȃ�����n��
�͑����ł��ł��Ȃ��Ƃ��Â��v���m�炳��Ă��鍡�����̍��ł���B�g��@���A����s���A�ɓ����v�Ƃ������i�H�j�u�`���E���v�̂����ꂽ���q�����w�҂���
������̂��A�q���̍�����ώ@���ʂ����낢��ƕ��́E�l�@���邭�������Ă������߂ł��낤�B����ł��u����DNA������j���[�X���^�[�v�ł́A������
�u�`���E���v�����ɍv���ł����̂ł͂Ȃ����ƂЂ����Ɏv���Ă���B
|
�ї��@�G�Y
�@
������w����ъ�b�����w�������@���_����
|
2013.05.20
No.12 �@�����ɂȂ�Ȃ��������ƍ����c�m�`������
���@�T��
�@�u�����v�́A���ɂƂ��Ă�������ƃR���v���b�N
�X�̓����
�������t�N�U�c�Ȋ������N���閂�@�̌��t�ł������B�q���̍��͍�������D���ł��������}�ӂ�����ĕ����u�����͍����w�҂ɂȂ�v�ƌ����Ă܂����
�����B�������A���w���̍��ɕ�e�Ɍ���ꂽ�ЂƂ��Ɓ\�\
�u���܂̍����w�҂͎E���܂���錤���������ĂȂ������߂Ƃ��Ȃ����v
�@���̈ꌾ�ŁA���͍����D���������w�҂̖����������Ă��܂����̂ł���i���Ȃ݂ɁA�����ƌ�ɂȂ��Ă��炻�̘b���ɂ����Ƃ���u���炻��Ȃ��ƌ�������
����v�Ƃ܂������o���Ă��Ȃ��悤�������j�B
�@�u�����v�ɕ��A����`�����X�����̌�Ɉ�x�����������B�k��ɓ��w�������́A�q���̍��̖����v���o���Ē����i����������j�̌˂�@�����̂ł���B����
���A�������������A�����������Â��i���X�ɘA��čs����Ēj�S�l�ŃR�[�q�[�������邾���̒n���Ȗ����i�����A�����͎����܂߂ĂS�l�������Ȃ������j�B
�u����ł́A���̐t�͏I���ȁv
�@�����v�������͑��X�ɑޕ����A�ȍ~�͒��Ƃ͖����̑�w�������𑗂邱�ƂƂȂ����B�������A�������Ɖ����������Ƃ����āA�Ƃ��Ɋy�����t���オ�K��
���킯�ł��Ȃ��A���͂����u�����v�Ƃ������t�ɑ��Ă��悢��[������ƃR���v���b�N�X���点���̂ł������B
�@�C�m�ے��܂Ŏ��͗L�@���w���w�B���̌�A�����w�ɓ]��������̊�b�����w�������ɐi�w�������A���m�ے��őI�e�[�}�͋��ނ̐��B�����w�������B��
�����A�����Ƃ͑S���W���Ȃ��B������A�ї��G�Y�搶�≪�c�ߐl�搶��啽�m�v�搶�Ƃ����X�S�������������߂��ɂ������Ƃɂ͂܂������C�����Ȃ���
���B�����ł��܂��u�����v�ɂȂ�`�����X���Ă����̂ł���B
�@���̌�A���É���w�Ɉڂ��Ă���A���������̌�������邱�ƂɂȂ����B���������Ȃ牽�ł��悩�����̂����A�����ăz�^�������C�t���[�N�̂ЂƂɑI��
�̂́A���Ԃ��ɑ��钷�N�̓���ƃR���v���b�N�X�̂����ł��낤�B
�@�R���c�L���V�ɂ������ނ�����Ƃ������ƂŁA�Ȃ�̏Љ���Ȃ��啽�m�v�搶�Ɏ莆�������Ă݂��B����ƁA�R���c�L���V�ɂ��đS���m���̂Ȃ���������
��ϐe�ɂ��w������������ɁA���V�ȎO�搶���Љ�ĉ�����A���傤�ǔ�����������̍���DNA������ւ̎Q�������߂Ă����������B���������Q������
�݂���A
�u����[�H�v
���B�����w�̕��ł͂悭�����グ�Ă����ї��搶��x���搶�������Ă����̂Ńr�b�N�������B���ӂ���Ƃ��B����������̂��i�B��Ă����킯�ł͂Ȃ��Ǝv��
���j�B
�@����DNA������́A���̂悤�Ȓ����R���v���b�N�X��������P�Ȃ郀�V�D����g�������Ԃɓ���Ă��ꂽ�B���ꂪ�������Ė���Q�����Ă��邤���ɁA���̌�
������䂪�z�[���O���E���h�ł���ł����S�n�̗ǂ��ꏊ�ł���悤�Ɏv���Ă����B����Ȗ�ŁA2010�N���{��̎����ǂ����������Ē������Ƃ�
�Ȃ����B�ق�Ƃ��́u�����v�ɂȂ�Ȃ����������ɁA���߂āu�����v�̊F����̂���`�����ł��邱�Ƃ����͂ƂĂ��������B
�@�{������ɂ́A���D�������̂܂ܐ��ƂƂ��������w�ҁA��������D��������ǂ����E�ƂƂ��Ȃ������Ȋw�ҁA��������Ƃ��Ă���A�}�`���A���D�ƁA����
�Ď��̂悤�ɒ����ɂ͂Ȃ�Ȃ��������������������Ă���Ȋw�҂ȂǁA���낢��ȃ^�C�v�̉�����������Ă���B�������A�S���̎v���͂ЂƂu�������D���v��
�̂ł���B�����Ă��݂����m�����܂��Ȃ����킹�Ă���̂��uDNA�v�̃L�[���[�h�ł͂Ȃ����Ǝv���i���̃L�[���[�h���Ȃ���A���̏o�開�͂Ȃ���
���j�B���̂��C�ɓ���̑f���炵�����̉�A���߂Ď��������ǂ𑱂��Ă������͑��������������̂ł���B
|
���@�T��
�@
���É���w��w�@�����_�w�����ȁ@���q�@�\���f�����O��������@����
|
2013.04.22
No.11 �@
�@ �R���� ����@��
��
�݁A���͕��Ɍ��O�c�s�̎O�c�s�L�n�x�m���R�w�K�Z���^�[�Ɋw�K�w�����i�O�c�s�����E���G�����S���j�Ƃ��ċΖ����Ă���B�W�����쐬�A���R�w�K�Ɋւ�闈��
�҃T�[�r�X�A���畁�y�����Ȃǂ���ȋƖ��ł���B���Z���^�[�̎�ȗ��َґw�͏��w���ȉ��̎q�ǂ��B�ł��邽�߁A�ł��邾��������₷�������́g�ʔ����h��
�`���邽�߂ɓ����Ђ˂���X�𑗂��Ă���i���Ȃ݂Ɏ��ȏЉ�ʐ^�Ɏʂ��Ă���͉̂ߋ��̓W���ɗp�����e�C�I�E�[�~�̕W�{
�ŁA���̊J���͂����ӂ��\�[�X�̗e
��Ɠ������炢�̑傫���ł��邱�Ƃɒ��ځj�B
�@�������̍D���̎q�ǂ������̂܂ܑ�l�ɂȂ����悤�Ȏ��́A�ޗnj��g��S���g�쑺�Ƃ����I�ɔ����̎R���Ő��܂������B2013�N
���݁A�l�����悻2000�l�̉ߑa�̑����B���̎��Ƌ߂��ɒ����\�����������Ă���A�������̋g
��n��͒J���[���n�`���}�s�ŁA�R�̎Ζʂɏ����ȏW�����_�X�ƕ��z���Ă���B21���I�ɓ���\���N
���o�߂������A���̎��Ƃł͂��܂��ɐd�ŕ��C���Ă���B�܂��A�Ăɂ͓����͖Ԍ˂����������J���������ɂ���̂ŁA�Ƃ̒��̌��ԂƂ������ԁA�Ⴆ�^��
�X�̈�����╨����̈����ˁA�_�I�Ȃǂ͎��I�̒��ԁA���Ƀn�G�g���O������郂���L�W�K�o�`�ɐ�̂���D�̑������������悤�ɂȂ�B���ɂ��삪���w
�Z�w�萅�j��ɂȂ��Ă��邱�Ƃ�A���ނ�̍Œ��ɖڂ̑O���J���l�Y�~���j���ł������̂Œǂ������ă^���Ԃł����������ƁA�ȈՐ����̎搅���ɃI�I�T���V��
�E�I���l�܂��Ă������ƁA�c�����X���b�p�𗚂����Ƃ����璆�Ƀ������K�������Ă������Ɓc���X�R���Ȃ�ł̘͂b�ɂ͖����ɂ��Ƃ܂��Ȃ��B
�@����Ȋ��ň�������͂ǂ������킯���A�c�������璎������ɋ���������A�ʉ��E�ʊw�̓����ʼn�����߂܂��Ă͌��ւ���̃v���X�`�b�N�P�[�X�ɓ���
��
��
�炵�Ă����B�c�t���̍��ɂ͑������u�ނ��͂����v�ƌĂ�A�l���ň�x�ڂ́g���m��(�ނ�)�h���擾�����B
�@�g��ƌ����X�M�̐A�т���ŁA�A���͂��܂�ʔ��������̂����A�W���̎��ӂɂ͏��K�͂ȃN�k�M�т��c���Ă���B���̎��Ƃׂ̗ɂ��N�k�M�т�����A����
����
�͂悭�g�Q���W�i�N���K�^�̂��Ɓj�h���̂�ɍs���Ă����B�ߋE�n���ōL��������n�������Ǝv���邪�A�g��ł̓N���K�^�̑��̂��Q���W�A�R�N���K�^
�̓w�C�P�A�m�R�M���N���K�^�̓X�C�M���E�A�~���}�N���K�^�̓}�N���ƌď̂��Ă����B
�@���w�Z1�N���̉āA���͈�l�ł����̃N�k�M�тɃQ���W���̂�ɍs���Ă����B��l���q�œ��N��̎q��
���̐������Ȃ��������Ƃ�����A�����̗V�ё���͒��������B�����Ă������悤�ɁA�ڐ���t�����N�k�M���R���ĉ���Ă�����E��Ɍ��ɂ��������B�X�Y
���o�`�Ɏh���ꂽ�̂��B���͂��܂�̋��|�ƒɂ݂ŋ�����߂��Ȃ���ƂɋA�����B�����̓L���J���őΏ����邭�炢�����Ȃ��A���̏����ȉE��̓p���p���Ɏ��
�オ�����B���炭�͎w���Ȃ��炸�������ĂȂ������̂ŃX�v�[���ł��т�H�ׂĂ����̂������Ă���B���̌ジ���ƁA���Z�����炢�܂Ńn�`�ɑ��鋰�|�S��
�Ȃ��Ȃ������邱�Ƃ͂Ȃ������B
�@���w�Z3�N�����炢�������Ǝv���A�Ƃ̒��Ŏ��]�Ԃɏ����K�����Ă����B��ɏq�ׂ��悤�ɕ��n����
���}�ȍ����Ȃ̂ŁA���]�Ԃ̗��K����悭�炢�����ł���ꏊ�����������B���x�������Ȃ���A���K���J��Ԃ��Ă���ƁA�O����傫�ȃn�`���������Ɍ�
�����Ĕ��ŗ����B���́u�A�J���I�h�����I�����ȁI�v�Əł��Ď��]�Ԃ̌�����ς��ĕK���œ����悤�Ƃ�����c�Ȃ�ƁA���]�Ԃ������ł���ł͂Ȃ����B��
�̎��A�n�`�ɒǂ��������Ȃ�������A���܂��Ɏ��͎��]�Ԃɏ��Ă��Ȃ���������Ȃ��i����Ȃ��Ƃ͂Ȃ��Ǝv�����ǁj�B
�O
��̃����[�R���������M���ꂽ�ē����q����͂��߁A���̒m�荇���̍����w�ҁA�����w�҂͓s��܂�̕��������B�����āA���ł������C��d�ŕ����Ă���Ƃ�
�����Ƃ͏o����������Ȃ��B��������ƍ�������ɑ��鋻���ɑ��ւ�����̂��ǂ����A���ɂ͕�����Ȃ��B���������l���䂪�傫�����Ă���̂�����A
���̊����Łg�������̍D���h����������̂ł���A�s��炿�̍����w�҂���w�҂������͓̂�����O���B
�@�O��̃R�����ŐG����Ă����ē����q����̋^��Ɠ��l�A����������̖ʔ����ċ����[�����Ƃ��q�ׂ�̂͗e�Ղ��̂����A�ł͉��̍D���Ȃ̂��A�Ƃ���
��
���ɂ͂�����l���Ă������邱�Ƃ��ł��Ȃ��B����͖{���ɓ�ł���B���ꂪ������Ύ����g�ɂ��Ă����Ɨ����ł���̂�������Ȃ��B�����Ă����ƍ����
��
�̖ʔ������q�ǂ������ɓ`���邱�Ƃ��ł��邩������Ȃ�
|
����@��
�@
�O�c�s�L�n�x�m���R�w�K�Z���^�[�@�w�K�w����
|
2013.03.20
No.10 �@
�@ �����يw�|���Ƃ��Ă��������� �ā@���@���@�q
����DNA������炱�̃R�����Ɏ��M�˗������������āA�܂Ƃ܂���DNA�̌������ʂ������͓̂���̂ŁA���������邩�ǂ��������܂������A�R����
�Ƃ������ƂȂ̂�DNA����O�ꂽ���ł����������������邩�Ǝv���A�����g�̂��Ƃ��������Ă����������Ƃɂ������܂����B�q���̍�����ǂ̂悤�ɒ��Ɋ�
����Ă������A�Ƃ������ƂƁA�������ق̊w�|���Ƃ��ē������œ��������Ă��邱�Ƃ����`�����Ă݂����Ǝv���܂��BDNA�̘b�肩�痣��Ă��܂��܂����A��
�����������������B
�@�����قō����S���̏����w�|���Ƃ��ē����Ă���ƂȂ��������D���Ȃ̂ł����H�ƕ�����邱�Ƃ��悭����܂��B�����Ă�����͓�ł��A�Ƃ��������Ă���
���B���e���Z�����ɐ������̂ɋ�������������ł��Ȃ��A������͓̂����̕i��Ƃ����A���Ɏ���ɒ�������������ł�����܂���ł����B����ł��Ȃ�
�����S�����������ŃZ�~�̂��������A�T���V���ɃA�Q�n�`���E�̗c����������Ɛ������y���݂Ɍ�������肵�Ă��܂����B���ł���I�I�X�J�V�o���n
�`���ƐM���āA�Ƃ̃K���X�˂̌��Ԃ���Ԃ��o���Ďh����Ȃ��悤�ɍ̂낤�Ƃ�����A�J�Ɍ����z�킹�Đ������R�b�v�ɓ���Ă����Ɛ��ʂɗ����Y�ށA�Ə�
���Ă���{�����āA�����̑������Ō����z�킹�����̂̋z����������Ă��܂������ƂȂǂ��A���w���̎��̓��Ɉ�ۂɎc���Ă���v���o�ł��B���̌�A���Ɏ���
�܂Œ��𑱂��Ă���ꂽ�̂́A���Ԃ�Ƒ��̋��͂ƕ��C�����������炾�Ǝv���܂��B���̂��߂ɒ|�̖_�ŕߒ��Ԃ̌q���Ƃ�����Ă��ꂽ�c���A�����V���`���E��
�H���̏u�Ԃ��ꏏ�Ɍ��Ă��ꂽ��Ƒc��A���e�Ɨ��s�ɏo�����Ă���ԁA�����Ă����A���W�S�N�ɃG�T�i�A���j������Ă��ꂽ�c��A�Ƃ������͎҂����܂����B
����ɁA�U���K�j�A�������A�w�r�̔����k�Ȃǂ܂ŁA���ȊO�ɂ��l�X�ȕ����Ƃ̒��Ɏ�������Ŏ��Ȃ��Ă������ɂ�������炸�A������������邱�Ƃ̂Ȃ�����
�Ƒ��́g���C�h�̂��A�ō�������ƁA��l�ɂȂ��Ă���C�Â��܂����B
�@�����̏��q���w�Z�ɓ��w���������l�Œ��𑱂��Ă��܂����B���w��N�̉ċx�݂̎��A���s�Ђ���悵�������̏W�̗��ɐe�q�ŎQ�����A���߂Ē��̓W���Ƃ���
���̂��K�����̂����������ɁA���̍̏W�ɂ͂܂�܂����B�u�V���������̏W�v���o�C�u���Ƃ��Ĕ�������O��q��Ȃǂ֍s���A�̂������Ƃ̖����`���E���̂���
�W�����邱�Ƃɖ����ɂȂ�܂����B�I�I�����T�L�����߂ăl�b�g�C���������̎�̐k���銴�������ł��ǂ��o���Ă��܂��B�̎����ɂȂ�ƁA���ɏ�����[��
�l���邱�Ƃ������A�Ƃɂ��������w�������̂����w�ɍs�������Ǝv���A���̌�܂��܂����ɂ̂߂荞�݂܂����B30�ōK�^�ɂ������قɐE�āA���݂܂�
�����ƒ��Ɗւ�邱�ƂɂȂ�܂����B
�@�����قł͎q���̎��R�����̑��k�������Ύ܂��B��������͎d�����Z�����̂ł��傤���A����Ă���͕̂�e�Ǝq���̂��Ƃ������ł��B�����Ă��q����
�قƂ�nj����J������e���肪����ׂ�܂��B�����Ďq���ɗ��h�Ȏ��R�����������������ǂ�����悢���A�Ƃ����ꍇ�������ł��B���̑��k���e�́A���Ƃ�
�A�����w�Z�֍s���O�ɉƂ̎���Œ��̃Z���T�X���q���ɂ��������A���N������������T�Ԃō��܂����A�ǂ�����悢���A�Ƃ��A�R���e�i�ɐ������r�I
�g�[�v������Ăǂ�Ȑ������̂����邩�ώ@���Ă������A�������N����Ȃ��̂Ń��_�J���Ă��ĕ���������ǂ���ς葼�ɉ����N����Ȃ��A���R�ώ@�ɉ���
�R�c�͂���̂ł��傤���A�ȂǂȂǂł��B�ċx�݂��c�肠�ƂP�T�ԂƂ��������ɂ���ė��āA�h��̎��R�����ɉ������Ηǂ����낤���A�Ƃ������k�������
���B������Ђǂ��o�����́A����ċx�ݎq�ǂ����k��ł̎��ł��B��e���q���̏h��Ə̂��ĎB�e�������̖��O�������ė~�����ƁA�R���p�N�g�f�W�J���̉t��
��ʂŎʐ^�̒������\�������肳�����Ă������̎��ł��B�ꏏ�ɗ��Ă����q���͂����ɖO���Ă��܂��A������ŃT���_���R����n�߁A���x�ڂ��ɏR��グ���T
���_�����ׂœ���Ή����������ʂ̐l�̕W�{���̒��Ɍ����ɗ������̂ł��B�������W�{�͉��A�����ł͂��܂���Ȃ��ƂȂ�܂����B��e���ʐ^�̒���
���O��t���邱�Ƃɖ����ł�������q���������̂��ƂȂ��������ł��B����Ȏ��͉��̂��߂Ɏd�������Ă���̂��킩��Ȃ��Ȃ�܂��B�����͋ɒ[�ȗ�ł�
���A�����ق֍����̑��k�ɗ���ꂽ���ꂳ��ɂ́A���������y�������Ƃ����܂��傤�A�ƌ����āA�قƂ�nj����J���Ԃ̖��������q���ɉ����D���Ȃ́H�ƕ���
�ƁA�N���K�^���D���A�A�����ʔ������A�Ȃǂƌ��\�������Ԃ��Ă�����̂ł��B
�@���̉Ƒ��͑O�q�̂悤�Ɏ����g���C�h���Ă����̂ŁA���h�Ȍ�������点�������甎���ق֘A��čs���A�Ƃ����悤�Ȃ��Ƃ�����܂���ł������A����ł���
�s�Ђ̍����̏W�̊��ɎQ�������Ă���܂������A���̌�̉Ƒ����s�͂������̌v�悵���̏W���s�ƂȂ�܂����B�܂莄�́A���e�́g���C�h�̌��ʁA�ʔ���
�Ɗ��������ɂ��Ă����ƒm�肽���Ȃ�A���ʂ̏��̎q���s�����������������w�����̂����w�ւ̓��w�������ꂸ�ɍ�������킯�ł��B�e�͎q����������
���������Ƃd���Ď菕�������Ă�����̂��ǂ��Ǝv���Ă��܂��B��������Β��ɂ͍����ɋ��������������A�����DNA�����ɓ���𖾂������ƍl����
��҂�����Ă����̂ł͂Ȃ����A���̂��������ƂȂ�Ɩ��N�ώ@��⍩���W�{�̂���������Ȃǂ𑱂��Ă��܂��B�����Ŏ��ۂɎq��Ă��������Ƃ��Ȃ���
�ł��܂�����͂͂Ȃ��A�q��Ă͂���Ȃɂ��܂������Ȃ��A�Ƃ�������邩���m��܂��A���ꂩ������炭�͊w�|���Ƃ��Ă����������𑱂��Ă���
���Ǝv���Ă��܂��B
|
�֓��@���q
�@
��t�������������� �w�|��
|
2013.02.03
No.9 �@
�@ �X�y�C���̎��R�ی�̈�[���_�Ԍ��� �Ɂ@���@���@�v
�ʐ^�́A�C�x���A�����i�X�y�C���j�̖k���吼�m�݂ɉ����ē�����300km�ɓn��̂т�J���^�u���A�R������Picos de
Europa���������̓��[�̋��E�̓��ɂ���n�C�C���O�}�̒����i�Ƃ��̂��ɗ����j�ł��B���̒����̓n�C�C���O�}�̕ی�‧���ƂɎ��g�ރJ���^�u
���A���i�����͂P�B�P���j�ɂ��ݒu���ꂽ���̂ł���Ƃ̂��Ƃł��B
�@���́A��N�i2012�N�j�X���ɃX�y�C���ŊJ�Â��ꂽ�u�v���X�~�h�����w���ۉ�c�v�ɎQ�����A�������ʂ̕����܂����B���̐܂Ƀs���l�[�R���t�߂ƃJ
���^�u���A�R���t�߂𗷍s���A�`���E�̍̏W�Ɗώ@������@��Ɍb�܂�܂����i�R�s���ɂ��ẮA���ĐM�B��w��w�@�Ńc�L�m���O�}�̐��Ԍ��������Ĕ��m
�̊w�ʂ����A���݃G�R�c�A�[�Ƃ���|���Ă���p���x�O�ݏZ�̃I�����_�l�Ɉē����Ă��炢�܂����j�B��L�̃n�C�C���O�}�̒����͂��̓r���ő����������̂�
���B����̗��s�́A���[���b�p�̃n�C�C���O�}�Ɋւ��钲���Ȃǂ̖ړI�ōs�����̂ł͑S������܂���ł������A���̋@��Ɉē����Ă��ꂽ�I�����_�l�̘b��C
���^�[�l�b�g�Œ��ׂ��X�y�C���̃n�C�C���O�}�̕ی�‧���Ƃɂ��ďЉ�����Ǝv���܂��B
�@���[���b�p�����̃n�C�C���O�}�͐�ł̊�@�ɒ��ʂ��Ă���A�t�����X�ƃX�y�C���̍����ɉ������s���l�[�R���ł́A1900�N�㏉���ɍŌ�̖쐶�̂�
�ˎE�����ł����Ƃ݂Ȃ���܂����B���̌�A���[���b�p�̑��n�悩��̎��Y�̂����x���ړ�����܂����B�������A���̒n��ŗL�̈���Ƃ݂Ȃ����̂���
���c���Ă����悤�ł����A�Ō�̌̂����N�O�Ɍ���ĎˎE����Ă��܂����Ƃ̂��Ƃł��i�ٔ������ɂȂ�܂������A�ˎE�����l�́u�g����邽�߁v�Ƃ̌�����
���F�߂�ꂽ�j�B�܂��A���̒n��͗r�⋍�̖q�{������ŁA�Ċ��͕��q���s���Ă��邽�߁A���Ȃ���ʔ�Q���o�Ă���A���Ƃ͊ȒP�ł͂Ȃ��悤�ł��B
�@�܂��A�X�y�C���̃J���^�u���A�R���̃n�C�C���O�}�ɂ��ẮA�J���^�u���A���ɐ�������M�������̓��ł͗B��n�C�C���O�}��1989�N�ȗ���Ŋ뜜��
�iendangered species�j�Ɏw�肳��A�ی�‧���Ƃ����{����Ă��܂��B
�@�X�y�C���̃n�C�C���O�}�̕ی�́A1973�N�Ƀn�C�C���O�}���܂މ���ނ��́u�쐶�����̕ی�v��ړI�Ƃ���@�������肳��A���̖@�Ɋ�Â���X�̋K��
�����ꂽ���Ƃ���n�܂�܂����B���̌�A1989�N�ɂ́u���R�n��Ɠ��A�����̕ۑS�v��ړI�Ƃ���@�������肳��A��Ŋ뜜��ɂ��Ă͉̂��߂̌v
������肷�邱�Ƃ����߂��܂����B�����ŁA���̖@���Ɋ�Â��Ă܂��n�C�C���O�}���܂ޑS���Ő�Ŋ뜜��J�^���O���Ҏ[����܂����B�ŋ߂ł́A2007�N
�Ɂu���R��Y�Ɛ������l���v�Ɋւ���@���ɂ��A���ʂȕی��K�v�Ƃ���쐶������̃��X�g�ƑS���Ő�Ŋ뜜��J�^���O���Ҏ[����A2008�N����̖@��
�ɂ��n�斈�̐�Ŋ뜜��J�^���O�Ҏ[�����߂��A�J���^�u���A���ł��n�C�C���O�}���܂ސ�Ŋ뜜��J�^���O���Ҏ[����܂����B
�@�J���^�u���A���ł́A1989�N�̖@���Ɋ�Â��Ē����Ɂu�n�C�C���O�}�v��v�����肳��܂����B���̌v��ł́A�J���^�u���A�R���S�̂̃n�C�C���O�}
�W�c�̈�`�I���l���Ǝ�Ƃ��Ă̐����������߂�̂ɕK�v�ȓ����̏W�c�Ԃ̌𗬂𑣐i���邱�ƂƂȂ�܂����B���̂��߁A�܂������̃R�A�ɂȂ�W�c�̕ی�A��
�B��}�邱�Ƃ�ړI�Ƃ��鎖�Ƃ��n�܂茻�݂Ɏ����Ă��܂��i�����͕ʂ̏B�A���ɑ����Ă��܂��j�B1989�N�̖@���ł́A�N�x���Ɏ��ƕƕK�v�Ȍv��ύX
��q�{•�_��•���Ƒ�b�i���A�q�{•����•�n��J����b�j�ɒ�o�����F�邱�ƂɂȂ��Ă��܂��B
�@2011�N�x�̕��i65�ŁA�J���[�j�ɂ́A���Y�N�x�̎�X�̃��j�^�����O�̃f�[�^�ƂƂ��ɂ���܂łɏW�ς��ꂽ�f�[�^�x�[�X�Ɋ�Â��܂Ƃ߂̐}�\
�i�n��ʂ̖ڌ����A�e�A��̎��O�}�̖ڌ����A�ƒ{�̔�Q�����̐��ڂȂǂȂǁj��ʐ^�i�����A�ߊl‧���b�̗l�q�Ȃǁj�������f�ڂ���Ă��܂��B�ō�30��
���[���i�I�j�̔����ƒ�߂��Ă���ɂ�������炸��������A�܂����̒n��ł��ƒ{��_�Y���ɑ����Q�͏��Ȃ��Ȃ��A��Q�ɑ���⏞���傫�Ȗ���
�Ȃ��Ă��܂��B�ʂ̎����ɂ��A�J���^�u���A�R���S�̂̃n�C�C���O�}�̌̐��͑�����140�����x�A�J���^�u���A���𒆐S�Ƃ��铌���̌Q��30���`
40���ɂ����Ȃ��Ƃ̂��Ƃł��B
�@�����m�̂悤�ɁA�X�y�C���͌��݂���߂Đ[���Ȍo�ϊ�@�Ɋׂ��Ă��܂��B���ł͂��̂��ƂɌ��y���āA�l�I�A���I�����̌����I�ȉ^�p�i�d���̉����
�ǁj�̕K�v���Ɛv���Ȕ�Q�⏞�̂��߂ɕی����x�𗘗p���邱�ƂȂǂ�����Ă��܂��B����ɂ��Ă��o�ϊ�@�̐[���ȃX�y�C���̈�n�������c�̂ł��ꂾ��
�̕ی�‧���Ƃ��p���A���{����Ă��邱�Ƃɂ͋�������܂����B�J���^�u���A���̓T���^�f�[���s�Ƃ����吼�m�݂̃��]�[�g�n�i�����A�M���̕ʑ��n���吼
�m��]�ދu�ɗ������сA�C�M���X�Ƃ̊Ԃɑ�^�D�ɂ�����t�F���[�q�H������j�����鏬���Ȓn��ł���A��r�I�o�Ϗ��u�܂��v�ł���Ƃ����w�i������
�����m��܂���B����ɁA�����Ȍ�Ɉ�x�͐X�т�O��I�Ƃ�������قǂ܂łɔj�����[���b�p�i���ɁA�����[���b�p�j�ł́A�i�ߖłڂ��I�Ɂj���ۑS�A
���R�ی삪���A�n�������c�̃��x���ł������ɍs���Ă���Ƃ����w�i�����肻���ł��B
�@���{�̖k�C���̃q�O�}�Ⓦ�k���璆���ɂ����Ẵc�L�m���O�}�ɂ��ẮA���Ȃ��Ƃ����ʂ͐�ł̊�@�ɂ͂Ȃ��A���R�̐H���A���̒��_�ɗ��N�}�Ɛl�Ƃ�
�����̂��߂ɕی�E�Ǘ����ׁX�Ȃ���s���Ɩ��Ԓc�̂Ȃǂ̋��͂̂��Ƃɐi�߂�����܂��B�������A��B�ł̓c�L�m���O�}�̊m���Ȗڌ��Ⴊ�₦�ċv��
���A�l���ł���ł̊�@�ɂ���܂��B�����̒n��ł͕ی�‧�v��̍����ڎw���đ��}�Ɍp���I�Ȍ̐������ƕی슈�������{����K�v������Ǝv����
���B
�@�Ƃ���ŁA�`���E�̍̏W�Ɋւ��ẮA�t�����X�ł͍��������A���R�ی�����Ȃǂ̊O�ł͋K��������܂��A�X�y�C���ł͖@���ɂ�肢���Ȃ��̃`���E��
�N�ł����Ă��i�X�y�C���l�ł����Ă��j�̏W���邱�Ƃ����̂��֎~����Ă��܂��B�X�y�C���l�̗F�l�i�����w�̌����ҁj�́A�����uNo one
cares.�v�ƌ����āA����������ǂ����Ƃ̈ӌ��ł����B�������A���p�̃g���u��������邽�߂ɁA�ނɎ�`���Ă�����č���̗��s�ŗ������\��̕���
�̌��ɌʂɃ`���E�̍̏W���\����O�����čs���܂����B�S���҂̍D�ӓI�ȑΉ��������āA����悭�R�����狖�邱�Ƃ��o���܂����B�s���l�[��J���^
�u���A�ŏo��������s�ҁA�n�C�J�[�A�r�����A�ѓ��Ǘ��҂̐l�������T���čD�ӓI�ŁA���l���̐l�����͎��̃��[���V�A�́i���R�j�`���E�̕��q�n���n���̌���
�̐����ɋ����������Ă���܂����B�����̐l���������S�ł������̂������ł������c�c..�B���Ăł̓t�B�[���h�Ń`���E��‧�̏W���Ă��ē��Ǝ҂ɏo��
�����Ƃ͂܂�����܂��A����������ł����B�����I�ȊW����X�y�C���̎R�Ŏq�������ɏo����Ƃ��Ȃ������̂͏����c�O�Ɏv���܂����B�u�`���E��߂�
�͈̂������Ƃ��v�Ƃ����悤�ȋ��炪�s���Ă��Ȃ���悢�Ǝv���܂��B
|
�ɓ��@���v
�M�B��w�@
���w�������Ȋw�ȁ@
���̐����w
���C�����@
|
2012.07.02
No.8 �@
�@ �����̖��͂�`����n���ȓw�͂��K�v�ł���B �h�@�q�@�d
�����̑��l���̎ړx�̈�͎퐔�ł���B����܂łɋL�ڂ���Ă��铮���̎퐔������ƁA�����͂��悻�V�����߂Ă���B������A�����͒n��������Ƃ����l
�����������Q�ł���A�n���͍����̘f���Ƃ������Ă���B������̍����ł��A�n��ɂ���Č`���������A�F�ʂ��قȂ����肵�Ĉق������邱�Ƃ��悭����B
���̂悤�ȑ��푽�l�������߂č����̏W�ɖ����ɂȂ鍩�����N�������B�������A�������N�̌����͊m���ɐi��ł���B���V�搶�A������Ɣ�������̃R����
�ł������Q���Ă���A���̌����Ɖ��P����q�ׂ��Ă���B�����������ΏۂƂ��Ă���l�ԂɂƂ��Ă͎₵������ł���B�������A����͎���̗���ł���
��A�����ȒP�ɂ��̗����ς��邱�Ƃ͓���Ǝv���B�ł��A�������N���������Ă��Ă��A�����̖��͂͌����Ă���킯�ł͂Ȃ��B����̂����鐶�����ɓK
�����A�l�X�Ȍ`�ԂƋ@�\�̑��l���𐋂��Ă���͍̂����ł���A�����̐i���E���l���Ɋւ��Ă͍��������ł͌�邱�Ƃ͂ł��Ȃ��B�n���ɍ����̖��͂𐢂̒���
�`���A�q�ǂ����҂������w�̌����Ɉ������邱�Ƃ��厖�ł���Ǝv���B
�@�����g���ŏ��Ɋ����������̖��͕͂ϑԂƋx���ł���B�𒎂Ɩђ������������X�ɂȂ�͉̂��Ƃ��_��I�ł���A�̂̑�ӂ��ŏ����ɗ}���A�H�ׂ����܂��Ɉ�
�������z�����Ƃ͔@���ɂ������������Ɖf��B������w�ō����x���@�\�̌����Ɍg����Ă����̂����˂Ă������������̖��͂������Ă�������ł��낤�B�I
�T���V�̌������n�߂�ƁA�����̌`�Ԃ̑��l���ɋ����䂩���悤�ɂȂ�A�����̏W���n�߂��B�q�ǂ��̍��A�������������Ƃ͂�����̂́A�����鍩�����N
�ɂ͉����y���A�I�T���V�͂������m��Ȃ������B��w�ł̌����͂����ς�g�D�A�זE�ADNA�Ƃ������~�N���̌����ŁA���������̂Ƃ��Ă������蒭
�߂����Ƃ��Ȃ������B�������A�I�T���V�̌����́A������Ƃ��̂��̂̓~�N���̐��E�ł͂�����̂́A���ʂƍl�@�͌́A�W�c�A��A�����͂���ȏ�̕��ތQ��
�ΏۂƂȂ�A�������̑S�̂�����悤�ɂȂ�A�����i���̌����ɑ��ݓ��ꂽ�B�I�T���V�̌������n�߂������̎��́A�n�����̌��������m�炸�Ɏ��s������d
�˂Ȃ���A�I�T���V�̌n���i���̌�������V�ȎO�搶�̂��ƂŐi�߂Ă������A�n�����̎}����������Ȃ���A�m���Ɍ`�Ԃ����Ă�����̂͌n������ɋ߂��W��
����A�t�͉����Ȃ邱�Ƃ�������ADNA�z��̏��ɐ����̐i���̗��j�����܂�Ă��邱�Ƃ��n�߂Ď����������͖{���Ɋ��������B�������A�S���͂����Ƃ͌�
�炸�A���炩�Ɍ`�����Ă���̂ɁA�n������ł͉����W�ɂȂ�A�t�ɖ����ɈقȂ�`�����Ă���̂ɁA�n������ł͎o���W�ɂȂ�A���������s�v�c�Ȍn����
��ꍇ�����X����B���̂��낤�B����̌n������`���ƁA�n��I�ɋ߂����̂��n������ł܂Ƃ܂邱�Ƃ͈�ʓI�ł���B�n��I�n�������̐������̂��A�܂�
���̒n��n���Ԃ̋��E���ǂ��ɂ���̂��A���������₢���o�Ă���B�v����ɁA�n�����������قNj^������������B�����������^����������ɂ͍X�Ȃ�
��͂��K�v�ł���A���R�ޗ��̒lj����W���K�v�ƂȂ�B���̂悤�Ɏ��̍����̏W�͕��q�n���̌�������n�܂����̂ł���B�̏W�͋�J���������A��J�����ߒ���
�o�ĕK�v�Ȃ��̂�����ꂽ�Ƃ��̖������͉����y�������̂ł���B�I�T���V�̌������n�߂Ĉȗ��A���N�t�B�[���h�ɏo�����āA���̂悤�ȍ̏W�̊y������
����Ă���B�����̏W���d�˂邱�Ƃɂ���āA�V���ȋ����[�����ۂ�������A�����̍X�Ȃ锭�W�ւƂȂ���B�����Ō��������̂́A�����̏W��ʂ��č�����
���͂����X�Ɋ�����邱�Ƃ�����A�����̂��������ō����̖��͂�m���Ă��獩���̏W���n�߁A���������ւƔ��W���邱�Ƃ�����Ƃ̂��Ƃł���B�q��������
���R�ɒ��ɐڂ���@��v�X���Ȃ��Ȃ�A���̎���ł͍����w�����҂̒n���ȓw�͂��v�X�d�v�ɂȂ��Ă���B
�@JT������������(BRH)�ł́A���N�̉ċx�ݒ��ɃT�}�[�X�N�[���Ƃ����C�x���g���s���Ă���B���̌������ł́A�g�߂ȍ������ޗ��ɂ��āA�Q���Ԃ�����
��DNA���o�APCR�ɂ��ړI��`�q�̑����A����z��A�n�����쐬�ƌ��ʔ��\�Ƃ�����A�̍�Ƃ��Q���҂ɑ̌����Ă�����Ă���B�u�����ڂɋ߂�����
���n������ɂ��߂��v�A�t�Ɂu�����ڂ��x����Ă͂����Ȃ��v�Ƃ�����|�̂��ƂŁA������̑I����s���ADNA�ɐ������̐i���̗��j�����܂�Ă��邱�Ƃ�
������Ɠ����ɁA�������@���ɕs�v�c�ŁA���͂̂��鐶�����̂ł��邱�Ƃ������ł���������Ă��炤�̂��ړI�ł���B�Q���҂ɂ͈ӊO�ɂ����w���������A����
�͍������N�����܂Ɍ����B�T�}�[�X�N�[�����Q���������ƁA�����ɋ����������A�����̏W���n�߂��q�ǂ�������B�����A�Q���҂̒����獩���w�����҂������
������������������Ƃ͂Ȃ��B
�@BRH�ł͂�����C�x���g��N�R��s���Ă���B����͎��������w�c�A�[�ł���B���ɂ̓c���O�������u���g���ēy�듮�����ώ@����v���O����������A�q
�������ɂ͏�ɐl�C�Ă���B���R�̗��t���̓y���̂��Ă��ēd�����g���ォ��M�������ēy���̏����������𗎂Ƃ��Č������Ŋώ@����B���̒��ł̈�ʓI
�펯�i�H�j�ł͌���ꂪ���̃��J�f�ނ�X�f�ނȂǂɑ��Ă��A�q�������͊��Ő^���Ɍ��Ă���l�q������ƁA���ݓI�ȍ������N�i�����j�͐̂������ς��
�����݂��Ă���Ɗ�����B���ɐڂ���@�������A�������������ݓI�ȍ������N�i�����j�͂����Ɩ{���̍������N�i�����j�ɂȂ��Ă����ƐM���Ă���B����
�������@����̒��ɑ�����A�₠�A��X�����̋@��𑝂₷�w�͂����Ȃ���Ȃ�Ȃ��̂��낤�B��������������N�̎��オ�܂��߂��Ă���̂����ł͂�
����������܂���B�����肢�����B
|
�h�@�q�d
�i�s�����������ف@
�n���i���������@��C������
|
2012.04.09
No.7 �@
�@ �f�����������̐V���㓞�� �V�@���@�P�@�K
|
�@���邳�烊���[�R�����̃o�g�����n���ꂽ�B���邳��Ƃ̍ŏ��̏o��́A2002�N�t�R�̐��Y�����ی𗬃Z���^�[�ōs��ꂽ�������[�N�V���b�v�̂�
���������Ǝv���B��ۓI�������̂́A���邳�p���������ޗ��̎a�V���ɂ���A����܂Ō������Ƃ̂Ȃ�������������J�Q���E�̈��́A���̋N���ɔ��邽��
�̊i�D�̍ޗ��ł������B���̌���l�X�ȋ@��ł��������铌�邳��̌����O���[�v���p���錤���ޗ��̃��j�[�N���ɂ́A�������₦�Ȃ��B�̏W�⎔��͍���Ȃ�
��������Ȃ����A�N�����������Ƃ̂Ȃ������������ł��邱�Ƃ͑f���炵���Ǝv���B�c��Ȏ퐔���ւ鍩�������������̑�햡�́A���ɑ��l���ɂ���Ǝv���B��
�̍����ɒ��ڂ���̂���������d�v�Ȍ��ƂȂ�B�Ⴆ�A�n����d�v�Ȉʒu�ɂ��鍩����f�������ł͖��炩�ɂ��邱�Ƃ̂ł��Ȃ������[�����ۂ���������
�ǂł���B����܂ł́A���Ƃ������[�����ۂ����݂��鍩���ł����Ă��ADNA�̌��t�ŗ������邱�Ƃ͍���ȏꍇ�����������A���̏͋}���ɂ�����
��B
�@1987
�N�ɔ��\���ꂽPCR�@�̕��y�ɂ��A�ǂ̐���������e�ՂɈ�`�q���������N���[�j���O�ł���悤�ɂȂ��ċv�����B���́A�ړI�Ƃ��錻�ۂ�������������
��`�q���ǂ�����Č����邩�ɂ���B�����̃Q�m�����ɑ��݂����`�q���́A�P��������x�ł���B���̗L���Ȉ�`�q�̒�����A�ړI�̈�`�q�������邽
�߂̃A�v���[�`�͗l�X�ł��邪�A�ǂ̕��@��p���Č����o�����͌����҂̘r�̌������ł���B�ŋ߂ł́A�R���s���[�^�[��ł̃N���[�j���O���\�ɂȂ����B
�S�Q�m�����̉�ǂ��s���v���W�F�N�g���i�s���Ă��邩�炾�B2000�N�̃L�C���V���E�W���E�o�G�̃Q�m����ǂ���ɁA�l�X�ȍ�����ŃQ�m����ǂ̕�
�������X�Ɣ��\�����悤�ɂȂ��Ă����B���̏�����ɉ���������e�N�m���W�[���o�ꂵ���B������V�[�P���T�[�ł���B1000�h���Ńq�g�̌l�Q�m��
����ǂ��鎞�オ�������Ȃ��A������V�[�P���T�[�̕��y�ɔ����A�{���̂��Ƃ��e�퍩���̃Q�m����͂��i�s���Ă���B�z��̃z�����W�[�Ɋ�Â��N���[
�j���O�̋�J���l����ƁA������1000�h���ō����̃Q�m���̊T�v����ǂł���Ƃ͊���������ł���B���M���ׂ��́Ai5k (5,000
Insect Genome
Project)�Ƃ���5,000��̍����̃Q�m����������5�N�Ԃʼn�ǂ���v���W�F�N�g�ł���B�����_�ł͂ǂ��܂Ō����������邩�͕s���ł��邪�A
�V���������Ɋ�Â����X����V�[�P���T�[�̊J�����i��ł��錻����ӂ݂�ƁA���̃v���W�F�N�g����������͉̂��������ł͂Ȃ��������B
�@
���݁A���f�������̑�\�͂����܂ł��Ȃ��L�C���V���E�W���E�o�G�ł���B���[�K���ɂ��1910�N�̔���ψّ̂̔����ȗ�100�N�ȏ�̗��j�����V���E
�W���E�o�G�����������́A�����̃��f���Ƃ�����萶���̊�{�������������������߂̃��f���Ƃ��Ĕ��W���Ă����B���f�������̒��̃��f���Ƃ��ĕs���̍����
�������闝�R�͑��X���邪�A��`�q�̋@�\��̓V�X�e���̖L�x���͈������B�����q�ɂ���`�q�g�����̂̍�o��1982�N�ɕ���Ĉȗ��A�l�X�ȉ��ǂ�V
�Z�p���������ꌻ�݂Ɏ����Ă���B����A�f�������̈�`�q�g�݊����̂̍�o��piggyBac�x�N�^�[�̊J���ɂ��傫���i�W�����B
1998�N�̃`�`���E�J�C�~�o�G�ł̐����ɑ����A���̂��Ƒo���ځA�����ځA�����ځA�����ځA�����ڂ̊e�퍩���ɂ����Ĉ�`�q�g�����̂̍�o������A��
���f�������ɂ����Ă���`�q�@�\��͋Z�p�����X�ɐi�W���Ă���B
�@
��`�q�̋@�\��͂ɂ�����v���́A1998�N�����Ŕ������ꂽRNA���iRNAi�j�ɂ���Ă����炳�ꂽ�Ƃ����Ă��ߌ��ł͂Ȃ��B��`�q�g�����̂̍�o
������ȍ����ł����Ă��ARNAi�@�ɂ��e�ՂɈ�`�q�̋@�\�j�Q���\�ɂȂ����B���̕��@�́A���S�ϑԍ�����s���S�ϑԍ��������łȂ����ϑԍ����̃V�~
�ɂ����Ă��L������������A�u���Ԃɕ��y�����B�������Ȃ���ARNAi�@�����\�ł͂Ȃ��͎̂c�O�ł���B��{��RNA��̍o�ɒ��˂������e��g�D�̍זE��
�Ɏ�荞�܂�Ȃ�����RNAi�������Ȃ��ꍇ��������ɂ���đ��݂���BRNAi�͖{���^�j�����̐����ɕK�{�̐��̖h��@�\�ł��邽�߁A�ǂ�ȍ����ł���
�E���ɓ�{��RNA�������Ă��܂���RNAi�͐�����B���ɁA�זE�����`�������ȑO�̑��j���E����̏�����ɓ�{���q�m�`�𒍎˂���ARNAi�ɂ��
��`�q�@�\�j�Q���ǂ�ȍ����ł�������ƍl������B���̂Ƃ���A������Ɉˑ�������{��RNA�̍זE���ւ̎�荞�݂̍��ق������郁�J�j�Y���͕s���ł�
��B���̌������𖾂����A�߂������ǂ̍����ł���`�q�̋@�\�j�Q���p�ӂɂȂ鎞�オ���邩������Ȃ��B
�@
�����ŋ߂̒��ڂ��ׂ��e�N�m���W�[��TALEN�i�s�������������������������@�`�����������������|�k�������@�d���������������@�m���������������j
�ƌĂ��l�H�k�N���A�[�[�ɂ��z����ٓI�Ȉ�`�q�^�[�Q�e�B���O�@�ł���B���̐�����́A�t�^�z�V�R�I���M�ƃJ�C�R��2012�N�̊w��ŕ��ꂽ�B
����A���̕��@�ɂ��f�������̈�`�q�@�\��͂��傢�ɐi�W���邱�Ƃ����҂����B
�@�V�����e�N�m���W�[�̊J���ɂ����@�_�I�ȏ�ǂ��ł��ӂ���A�u���[�N�X���[��
�Ȃ錤���ւ̔��W��������Ă����B���f�������Ɣf���������킯��Z�p�I�Ȋ_���͂܂��܂��Ⴍ�Ȃ��Ă���悤�Ɋ�����B����̍����c�m�`�����̔��W���y
���݂ł���B
|
�V���@�P�K
���É���w��w�@�����_�w�����ȁ@���������Ȋw��
����
|
2011.11.22
No.�U �@
�@ ��r�����w���番�q�n���n���̐��E�� ���@��@�K�@��
|
�@�u�����{��k�Ёv�̘b��i�i������j����u��`�q����̕W�{�Ǘ��p�v
�i�q������j�ցA�����Ď��ւƃR�����������[����Ă����B�����g�A�Ôg��Q�ɂ��n��̌Q��ł̉\���������Ƃ����A�Ƃ���N���������̈�`�q��͂�
�ւ�邱�ƂƂȂ����B
�@��
�̌̌Q�́A������ꌴ�q�͔��d���t�߂̂��߁A����c������s�\�Ȃ̂ł��邪�A�k�БO�ɕW�{���m�ۂ���Ă������߂ɁA���̂悤�Ȉ�`�q��͂̋@�����
�ꂽ�̂ł���B�ߗׂ̌̌Q����͒n���I�ɂ��Ȃ�u������Ă���A���������ł͂�����̌̌Q�ł��������ƁA���A�Ôg�̉e������قǂɊC�݂ɋ߂�
�ȂǁA���Ȃ���قȌ̌Q�ł��������炵���B����̐k�ЂƒÔg����X�̑z�������̂ł������Ƃ��Ă��A��N�E����N�Ɉ�x���炢�̃X�p���œx�X�N����
�Ă����悤�Ȏ��R���ۂł���ɂ͈Ⴂ�Ȃ��B�����ł���Ȃ�A�ߋ��ɂ��A�Ôg�ɂ��h���ɎN����Ȃ�����������Ă����̌Q�Ȃ̂ł́H�@�Ƃ����ӌ����ł�
���悤�B����������̏ꍇ�A�l�Ԋ����̉e���ɂ��̌Q�T�C�Y�̏k����������ꂽ��ɁA��K�͊h�����Ă���̂ŁA����܂ł̂��̂Ƃ͈Ӗ�������Ă���
�̂��낤�B
�@�����炭���k�n���̑����m���݂ɂ́A�����悤�ȋ����ɂ��鍩����Q�����ɂ�����
���̂Ǝv����B���̂悤�ȏ��ɂ����āA�u���̏ꏊ�ɁA���̍���������
�Ă����v�Ƃ��ĕW�{���㐢�Ɏc�����Ƃ�A��`�q���̂悤�ɕW�{����܂�����������āA������ƃA�[�J�C�u���Ă������Ƃ͏d�v�ł��낤�B���͂Ȃ���
���A���̂悤�Ȍ`�ł̍v�����ł���ƍl���Ă���B
�@�����ނ��܂ނ����鐶����Q�̈�`�q���́AGenBank��
��ы�����������Ă���f�[�^�x�[�X�ɏW�ς���Ă��邽�߁A�����ɓo�^���ꂽ�f�[�^�Ɋւ��Ă͖��l�����p�\�ƂȂ��Ă���B�����A���{�ɂ͐������Ă���
���悤�ȍ�����Q�̔z��f�[�^�Ȃǂ��d�ė��p�����Ă��������Ă���B�莝���̃f�[�^��GenBank��
��擾�����f�[�^�������Čn����͂��邱�Ƃ������̂����A���X�A�s���Ȍ��ʂ�����������邱�Ƃ�����B���̈���ɂ́A�o�^���ꂽ��`���ƁA���̈�`��
��̂�����ł������Ƃ��ēo�^����Ă���햼�̑Ή�����v���Ă��Ȃ����߂̂��Ƃ������悤�ł���B�ނ��A�ڂ�ȃ��x���Ƃ����������n����͂Ɏ���������
���錤���҂ɂƂ��ẮA�탌�x���̓��萸�x���������ʂ����E���邱�Ƃɂ͂Ȃ�Ȃ����߂ɁA����قNj����S�ɂ͂Ȃ�Ȃ��ł��낤���A�ł�����莯�ʂ̗e��
�Ȏ��ΏۂƂ���悤�Ȋϓ_����̃^�N�\���E�T���v�����O���Ȃ���Ă͂��邾�낤�B
�@�������Ȃ���A�����ΏۂƂ��Ă��鐅�������ނł́A��̑q������̃R�����ŏЉ�
���ꂽ�u�����ނ��v�̗�̂悤�ɁA������u���ʎ�v�Ə̂���Ă���悤��
��ɂ����Ă������B����̑��݂����炩�ƂȂ����肵�Ă���̂�����ł���B�b���ނ⒱�ނƈ���āA�}�C�i�[�ȍ�����ΏۂƂ��Ă��邪�̂̈ӌ��Ȃ̂�������
�Ȃ����A�q�����q�ׂĂ���悤�ȁu��`�q���o�ɗp����ꂽ�W�{�́A���ł����ފw�I�Č������\�ȏ�Ԃŕۑ������ׂ��ł���v���Ƃ�Ɋ����Ă���B
����ł��������X�y�[�X��肪���܂Ƃ����{�̔����َ�����l����ƁA�����I�ɂ͓�����ƂȂ̂����m��Ȃ����A�핪������舵����������L�ڂɊ֘A����
�����ɂ����āA��`�q��p����ꂽ�ꍇ�Ȃǂɂ����Ă͓��ɁA���҂̐Ӗ��Ƃ��Ĉ�`�q���o�ɗp�����W�{�ނ�R��ׂ��@�ւɎ��߁A�W�{�̏��݂�_�����ɂ�
���L���ė~�������̂ł���B�������邱�Ƃ�����ꍇ�ł��A���߂āA�̏W������Y�n��_�����ɏڂ����L����Ă���Č������Ȃ����ۂɂ͖��ɗ���
�ł͂Ȃ����낤���B
�@�����܂ŁA�k�Ђ̂��Ƃ��`�q��͂ɋ����ꂽ�W�{�̈����Ɋւ��鎄��
���q�ׂĂ������A�㔼�ł́A�������g�̍���DNA�����ɂ��ďЉ���Ă������������B
�@���̍��������̂͂��܂�́A�u��r�����w�v�Ə̂���镪��ŁA�����̑̂Â����
�ʂ��āA���l�ȍ����`�Ԃ̊�{�v�����𖾂炩�ɂ������A�Ƃ����Ƃ���ɂ���
���B�Ȃ��ł��A�������ł����n�I�ȍ����ނł���J�Q���E�ނɒ��ڂ��A���������Ђ�����`�Ԋw�I�ɁA�����đg�D�w�I�ɒNj����Ă����B�w�ʎ擾�܂ł�
DNA�Ƃ͖����ɉ߂����Ă������̂́A������`�w�I�������A�V���E�W���E�o�G���͂��߁A�J�C�R�K��R�N�k�X�g���h�L�ȂǂƂ��������f�������ނ���A�n���i
���̌���
����L�[�^�N�T�ւƏ��X�Ɋg����͂��߂����ł����������߁A�L�������ނ̐i���̌�������J�Q���E�ނł�������`�w�I�����ɊS�������͂��߂�悤�ɂȂ�
���B�������A���̎�̌�����@�ɂ͑S���̑f�l�ł��������߁A�l�X�ȕ��X�Ƃ̋���������ʂ��āA�J�Q���E�ނɂ����锭����`�w�I�Ȍ����ɂ�����˂����ނ���
�ƂȂ����B
�@���̂悤�Ȓ��A7�N���قǑO�ɁA���݂̐M�B��w���w�������Ȋw�Ȃɒ��C��
���B
�ꂩ�猤�����𗧂��グ�Ȃ���Ȃ�Ȃ��ɂ��������A��`�q��͂Ɋւ��ẮA���Ȃ�̕������w�ȋ��ʋ@��ނ𗘗p���邱�ƂŃJ�o�[�ł������߁A�T�[�}
���T�C�N���[�≓�S�@�A�①�E�Ⓚ�ɂ₻�̑��̏����ނ��������Ă��܂��Β����Ɍ������X�^�[�g��
���闘�_���������B�����āA�L���Ȏ��R�����͔����ɑ��݂���M�B�̒n�̗��͉��������肪�����A����܂ł͑S���o���̂Ȃ��������q�n���n���w�I�Ȍ����ɂ�
�V�K���肷�邱�Ƃ������B����ɍK�^�������̂́A��w���Ɂu�R�x�Ȋw�����������v�Ƃ����g�D�������オ��A���̌������̌��C�ɂ��Ă������������Ƃł�
��B���{�A���v�X�n��𒆐S�ɁA�R�x�Ȋw�ɊW����l�X�ȏ�W�ς��A�t�B�[���h�ł̒����E�����ɂ��X��}���Ă���������Ƃ����A���̏�Ȃ�������
�������Đ������̂ł���B
�@
���ɒ��肵���̂́A��l�I�O���i��180
-100���N�O�j�̑��䍂�A��̎R�x�`���i���N�j�ɂ���ĉ���������ɂȂ����Ǝv���錴�n�I�J�Q���E��Q�Ɋւ��錤���ł������B���ۂɈ�`�q��͂�����
�݂��Ƃ�
��A�����ɑ��䍂�A��̓����i���{���Ɣ�ˑ��j�ň�`�I�����������Ă��邱�Ƃ����炩�ƂȂ����B�����āA���肳�ꂽ���R�[�̌̌Q�Ԃł̕���N����n�j��
�悭���v�����
�̂ł������B�܂��A���̎R��ł́A�Ċx�ΎR�Q�̊������ɔ����A�͐�̍ǎ~�߂Ɨ��H�ύX���p�ɂɐ����Ă����Ƃ�����邪�A���̂悤�ȁu�͐쑈�D�v���ۂɊւ�
�Ă��A��`�q��͂̌��ʂ͋����[���֘A���������Ă��ꂽ�B
�@���̂悤�ȓW�J�����������ɁA�R�x��ɂ����镪�U�͂̎ア������Q�ɒ��ڂ��邱
�Ƃ͓��P���Ȃ�����A��͑ΏۂƂ��鍩���Q���g���Ă����B�ŋ߂ł́A�Ώۂ�
�n����g���āA�쐼�������܂߂����{�L��A����ɂ͒��N������嗤�Ȃǂɂ������^�Ԃ悤�ɂȂ��Ă����B���̂悤�Ȍ������͂��߂������A���݂̕��z����
�ʂ������̂ł���A�����悤�Ȑi���j��H���Ă����̂��낤�ƍl���Ă����̂����A���ۂɏڂ�����͂����Ă݂�ƁA�����P���Ȃ��Ƃ���ł͂Ȃ����Ƃ��悭
�������Ă����B�ЂƂ��Ɂu���{�̍����̋N���⍩�����̐����v���Z�X�v�Ƃ����Ă��A���ɑ��l�ȃp�^�[����������̂��ƁA�����A�y���݁A�����ĕs�v�c����
��Ɋ����Ȃ���߂����Ă���B
|
����@�K��
�M�B��w���w�������w��
����
�i���C�@�M�B��w�R�x�Ȋw�����������j
|
2011.10.24
No.5 �@
�@ ��`�q����̕W�{�Ǘ��p �q�@���@�ǁ@��
|
�@�w��
���ނ��x�Ƃ��鍩��������B�w�����ނ��x�́A���������Ɨ�����̐��ɐ��ސ��������i�c���j�̑��̂ŁA�M�B�Łw�����ނ��x�͗L���ȐH�ނł���B�w����
�ނ��̒ώρx�Ɏg���l�X�ɂ��Ȃ��݂̐[���q�Q�i�K�J���g�r�P���̈�`�q���e�n�ō̏W���Ē��ׂĂ݂�ƁA����܂Ńq�Q�i�K�J���g�r�P���Ɠ��肳��Ă�����
�̂̒��ɕ����̎킪�������Ă������Ƃ����炩�ƂȂ����B���������q�Q�i�K�J���g�r�P���̋߉���́A�q�Q�i�K�J���g�r�P�������܂�ɕ��ʎ�Ȍ̂Ɍ���
����Ă����̂�������Ȃ��B
�@��`�q�Ŏ��ʂ��ꂽ������c������A��̔F���ɕK�v�ȊO���`�Ԃ̏�W�܂�A
���{�̉͐쐶�Ԍn���\����q�Q�i�K�J���g�r�P���̋߉��킪�V�����^�N�T��
���Ēlj�����悤�Ƃ��Ă���B������`�q����͂����̂��ǂ̌̂�������Ȃ��悤�ȏ�ԁA���Ȃ킿�W�{�̐������[���ɂ���Ă��Ȃ������Ȃ�A���̌�����
�����ł��Ȃ������ɂ������Ȃ��B��`�q��͂��ꂽ�̂�����ł��āA������`�ԉ�͂ŗ��ł��ĐV�����^�N�T���F���ł����̂ł���B
�@��`�q���i����z��j�́A���ތQ���i�����̊w���j�ƌ��т����Ă���B����
��`�q���͂��������̓���Ɍ�肪����A���͂����������ǂ̌̂���
��ł������ؕs�\�ȏꍇ�A���̏���p�������ʂ̉��߂ɑ傫�ȉe�����o��͎̂����̗��ł���B���̈Ӗ��ň�`�q��͏؋��W�{�́A���ފw�ɂ�����w�^�C�v
�W�{�x���Ȃ킿��̒�`�Ɏg��ꂽ�W�{�ɏ����鈵������ׂ��ł���B
�@��`�q��͂��ꂽ�����W�{�́A�ǂ̂悤�ɕۑ��Ǘ�����Ă���ł��낤���H��ʂ�
��`�q��͂Ɏg����W�{�́A�A���R�[���t�Z�W�{���������܂��܂Ȍ`��̕r
�ɓ����Ă��邱�Ƃ������B����䂦�i�z�˂���j�Ǘ�����ϔώG�ł���B�W�{�̕ۑ��ɂ��Č����̌���͑�ϋ�J�����Ă���B��J�̍ő�̌����͎��[�X
�y�[�X���[���m�ۂ���Ȃ����Ƃɂ����̂��B�i�����W�{�́A���x�ɋK�i�����ꂽ�w�h�C�c���x�Ƃ��錘�S�Ȗؐ��̔��Ɏ��[����t�Z�W�{�Ɋr�ׂĊǗ�����
�₷���j�B���x���ɏ������ޏ��͂��Ƃ��A���x���̍ގ���L���Ɏg���C���N�Ȃǂ̑I�����ێ��Ǘ��ɂ͏d�v�ȗv�f�ł���B
�@��`�q���o�Ɏg�����W�{�͂�
�ł����ފw�I�Č������\�ȏ�Ԃŕۑ������K�v������B�W�{�̈ێ��Ǘ��́A�����Ė����łł���悤�Ȃ��̂ł͂Ȃ��l�����
�܂߂��o��K�v�ł���B�����̊����W�{�̏ꍇ�A�����ق��l����W�{��a����A�W�{���ɔz��E�f�[�^�x�[�X�ւ̓o�^�E���J�ɂ�����o����Օi�Ɛl�����
�܂߂ĂP�̂������T�h���Ƃ��Ă��镶�������������A�t�Z�̈�`�q��͏؋��W�{�̏ꍇ�K�v�Ȍo���1�̂�����1500�~������Ȃ��ł��낤�B����͎�
���������Ă��鐅�������̏ꍇ�ł���A���ׂĂ̍����ł��ꂾ��������ƌ����̂ł͂Ȃ��B���\�o���������̂ł��邪�����̍̏W�⓯��ɂ����������Ԃ�J
�́A��`�q��͂ɂ�����
���o��A�����̌��؉\����S�ۂ���Ƃ������Ƃ����킹��Δ[���̂ł��Ȃ����z�ł͂Ȃ��B
�@�Ő�[�̂��Ƃ���ɖڂ������Ă���ƁA�v��
�ʏ��ɗ��Ƃ���������ł���B�������J�����đf���������������Ă��A�����؋��W�{���Ȃ��ƐM�p����Ȃ�����
�ł���B��`�q��͏؋��W�{�̖��́A���R�������������n�Ŕ����ق𗈊َҐ��ł����]���ł��Ȃ��悤�Ȍ����邩��S�z�ɂȂ�̂ł���B�W�{���������_
��Z�p�͊m���ɌÓT�I�Ȃ��̂ł͂��邪�A�Ő�[�̂������낢���������̂悤�Ȋ�b�̏�ɐ��藧���Ă��邱�Ƃ����̋@��Ɏv���N�����Ă���������K���ł�
��B
|
�q���@�Lj�
��t��������������
��Ȍ�����
|
2011.9.22
No.4 �@
�@�@�����{��k�Ђ���w�Ԃ��� �i�@���@�Á@�V
2011
�N8��7���A�����m���݂𑖂��鍇�Ԃ�D���āA�R�`�����̎R�ŕ�����T�������Ă����B30�x���鏋�����ŁA�G�]�[�~�̑升���̃X�M�т�������ʂ��A
�������\�N���ʂ�l���Ȃ��܂܂Ɏ���ꂽ�������x�������Ȃ���A�悤�₭������̌E�n���݂��ĉ����Ă݂��B�ۂނ����̊Ԃ��琁���o����C�ɁA�����}��
��₳��ĐS�n�悢�B�����ɂ́A����Ă͂�����̂̌����ȐΑg�݂��c���Ă����B���āA�����͋ߋ���~�̃J�C�R�̗��̒����ɂ������B�₽���̏�ɐQ�]��
�Ȃ���A�X�M�̏��𗬂��_�̍s����ǂ��Ă����B
���̉ẮA�Â�����̗{�\�𑱂���
�_�Ƃɗ��݂���ŁA�����Ԋu�ŁA�d���̗l�q���B�e�����Ă��������Ă����B94��70��
��l�̒j��ɂ�鉊�V���̃N������A�\���t��H�ׂ鉹�B�R�`���암�ł́A���łɗ{�\�_�Ƃ͂��̂P�������ɂȂ��Ă����B���̉Ƃ������A���ĕ����ɗV�т�
�s���ĐΎ��̒�������@�艺���A�X
�̉���r�߂ėV�Ƃ����b�����Ă����������B��C�̕����o�����ł͈�N���C�������肵�A�J�C�R�̉H�������߂ł���B�����̐Ύ��́A�l�Ԃ��C����
�Ǘ��ł��Ȃ���������́A�����̒m�b�̌����ł��������B
�①�ɂƂ������̂��o�ꂵ�Ă���A
������K�i���Ƃȁj���l���Ȃ��Ȃ�A�����I�̊ԂɐΑg�͕��ꂩ���āA�N���d����}�u�h�E
�̖��ɕ����Ă����B
3��11���̓����{
��k�Ђ̂��ƁA���R�j�����̔�Q�͂����Θb��ɏ�����B���O���c�̔����ق̕W�{�����ɑ����̐l�X���ւ�������Ƃ͗l�X�Ȕ}�̂ŕ�ꂽ���߁A�l�X�̋L
���ɐV�������낤�B
�c�m�`�����̗̈�ł́A�ЊQ���̎����ۊǂ͂ǂ����낤���B�ቷ�ŕۊǂ�
�˂Ȃ�Ȃ����̂��啔�����B��K�͒�d�Ƃ�����肪������ƁA�①�{�݂��@�\���Ȃ��Ȃ�B�c��ȗⓀ�������A�S���e�n�ɎU��镗���ɒS���グ�A�Ύ���
�[�����Ē�������킯�ɂ͂����Ȃ��B
�@
�܂��́A�a�@���w���̂悤�ɁA���p�d���ւ̐�ւ����\�Ȉ��肵���ۊǎ{�݂̊m�ۂ��O��ɂȂ�B�V�[�P���T�[���͂��߂Ƃ��������{�݂⎎��́A����
�̒��K�v�ɂȂ��ʂ����邩������Ȃ��B�Ɩ��p�Ⓚ�ɂ��������Ƃ��Ă��A���p�d�����S���ƂȂ��{�݂ł́A�ĂȂ�ΐ����̒�d�ł��n�����Ă��܂��B��
��ɁA��K�͂ȉЂȂǂ������₷���H�ƒn�сA���邢�͔R�������{�݂⌴�q�͎{�݂̎��͂Ȃǂ��A�ЊQ���ɉ����N���肤�邩�Ƃ������Ƃ�O���ɒu���Ă�����
�Ȃ�Ȃ����낤�B
�k
�В���ɁA���炭�g�����̎��Ȃ��������}������߂�Ƃ��A�����͎��͂̎d���̎ז��������Ă��Ȃ������A�����̖@��w�����ɍv���ł����ʂ͂Ȃ���
�̂��ƁA�Ȃ��m�b���i�낤�Ƃ����B�A�������ׂēr�₦���Ȃ��ŁA�c�m�`��͂̎���𓌋�����R�`�ɉ^�Ԃ��Ƃ͂ł��Ȃ����B�c�m�`��͂ɂ��l���ʂ̎��v
���}�����������Ƃ̎v�����������B���v���A�{���ɕK�v�Ȃ��͎̂��q���ً̋}�A���Ԃł����ɔ��������̂����A�����͂����܂œ������Ȃ������B�������A
����̉^������������Ȃ������B�Ⓚ�ł̑�z�ւ��ғ����Ă��Ȃ���ԂŁA�������̃h���C�A�C�X�͔�Вn�̈��u���ɑ����A���Ԃɉ���̂͂Ȃ���������
���B���ǁA���͋����̏�i�̕��X�ɐS�z�������������ŁA���i���ɗ����Ȃ��l�Ԃ͔�펞�ɂ����ɗ����Ȃ��A�Ƃ������Ƃ��ؖ����������ɏI������B
��
��������ʂł̊�@�Ǘ����A��������Ƃł��Ă��邱�Ƃ����Â����S�����̂��V���Ђ������B�H��̑傫�ȗ֓]�@�́A�d���̑���Ȃ����p�d���ł͓������A
�R�`�V���Ђł͏H�c�E�V���E�����Ȃǂ̊e�ЂƘA�����Ƃ�A���ʓI�ɐV���ň�����ċ}�����A�����̔z�B�ɊԂɍ��킹���Ƃ����B��d�������ăe���r�E�p�\�R��
�̎g�p���������Ԃł́A�W�ʂ̗Վ��łƂ͂����A�V�����l�X�ɉ^�傫�Ȏʐ^�̏��͏d�������B
�����ɒm�b����g���ė����̒������Ԃɍ��킹�����A�Ƃ����b���A�R�`�s
���̈��݉��ŎR�`�V���Ђ̃f�X�N���畷���Ȃ���r�[���̃W���b�L���X���Ă����̂́A4��7���̐[��̂��Ƃ������B���̒���A�ő�̗]�k���N�����ēX�̂Ȃ����^��
�ÂɂȂ�A�����Ɉ��͂��̃f�X�N�̕��X���u��d�ŗ֓]�@���~�܂�B���Ԃ��x������A�O��i3��11���j�����܂������I�v�ƌ��t�����킵�Ȃ���{�ЂւƋ}���ꂽ�B������
������8��30���A�V���ň�����ꂽ4�ʂ����̒���
���R�`�����̊e�˂ɓ͂���ꂽ�̂��B
���̐k�Ђ̂Ȃ��ɂ����āA���O�̖��S�ȑ�ɂ���Č�������������V��
�Ђ́A��@�Ǘ����ł��i��ł����g�D�ɈႢ�Ȃ��̂����A�@�ւ����Ƀj���[�X�ɂ��L���ɂ��o���A�ӊO�Ȃقǐl�X�ɒm���Ă��Ȃ��B
�@
��Ў҂͊F�A�u���ꂽ����A���ʂƂ��ċ��x�Ȑ��_�͂ŏ��邱�Ƃ��������A����ꂽ�W�{�̂��Ƃ��Ă��\���������Ɂu�ЊQ�ł�����v�ƌ����
��B�Ί���₳���A�����Ďx���ɑ��銴�ӂ������ɂ��Ȃ��B�ނ�̌�����{�����o�邱�Ƃ́A�����͂Ȃ����낤�B�����炱���A�u�x�������v
�u�������ꂽ�v�Ƃ����A�O���̎�������̔��k�ŏI����Ă��܂������ɂȂ�B
�Ƃ���ŁA�~���̐����オ�������I�����ق̕W�{�ɂ͑Ƃ�ꂽ���A�����ɏ���
����ʌl�����̉���A���̕W�{�A�ʐ^�A�m�[�g�A�������A�ƂƂƂ��ɊC��
���������ƂɒN���G�ꂸ�A�����ًƊE�̑g�D�I�x�����������グ���邱�ƂɁA���ǂ������������邱�Ƃ��m�����B���{�̎��R�j�����́A�قڌl�����̒~��
�ɂ���Ďx�����Ă������Ƃ͘_��҂��Ȃ��B�����ق̕W�{���R��B����̂悤�Ȕ�펞�ɁA�����������Ԃ̌l�����́A�c�O�Ȃ��犮�S�ɉᒠ�̊O�ɒu�����
�����B�ǂ̂悤�Ȕ�Q���������̂��Ƃ������̈ꌳ�������Ȃ���Ă��Ȃ����Ƃ́A���̓��{�̎Љ�̑傫�ȗ��Ƃ����ł�����B
�Ⴆ�A�����c�m�`������ŁA�Ⓚ���ꂽ�c�m�`�����̕ۊǏƁA�ЊQ���ɑz�肳
���댯���ɂ��ď����Ƃ�܂Ƃ߁A���ꂼ��̎{�ݓ��ł̑�Ɍ��т�
�Ă͂ǂ����낤�B�ۑ�̋��L�Ȃ����ẮA����������悤�Ȃ��Ƃ��J��Ԃ����B�������A�����ق̎x���������̓w�͂ɂ���Ďx�����Ă��邱�Ƃ́A��[��
�ւ�������̂Ƃ��Ă悭���m���Ă���B�����A���ꂾ������k�ɏI��点�Ă��܂��ẮA�W�{��m�[�g�ƂƂ��ɏ������ݖ�̌����҂̏�M���A���܂ł�������Ȃ��̂��B
|
�i���@�ÔV
���R�ʐ^��
|
2011.8.10
No.3 �@
�@�@��s�����i�K�T�L�A�Q�n����������� ���@���@���@��
|
�g�߂Ȑ��������̏W���Ė��O�ׁA��������������ώ@���邱�Ƃ���
�����w�͎n�܂�|�|�������̃x�r�[�u�[�}�[����́A������������߂Đ^�����ȉȊw������Ĉ�����B�����̏W�����コ��A�W�߂��W�{�����̂܂�
�Q����A���ꂪ�ċx�݂̏h��ɂȂ�B���R�����̃e�[�}�ɔY�܂���邱�ƂȂǂȂ��A�������N�ɂ͂��肪��������B�������̂ł���q���A�����������̂�
�Ă����̕����A�q�������ł̏���͂����ƍ��������B�����琢�̒��͕ς���Ă��܂����̂��c�c�B
�������N�̐��͌��݂̔�ł͂Ȃ��A�`���E�𒆐S�ɁA���Ɛ��A���邢�͓�Ɩk�̎q��
���ʂ�ʂ��čs���W�{�̌������͂�����B�G���u�q���̉Ȋw�v�������낤���A��]�̎�ƒł������L���ĕ��ʑ�������߂�R�[�i�[���������B
��������A���̃l�b�g���[�N�ɉ�������B�֓��A�����̎q��������
������A�����A�썑�E�F�{�s�ɏZ��ł������ɂ͂��Ȃ�L���������B��̒��ɂ̓~�J�h�A�Q�n�A�N���R�����T�L�A�T�c�}�V�W�~�ȂLj�H�v���Ȃ��Ɠ�����
�����̂����������A�u��͏��i�v�̃i�K�T�L�A�Q�n��c�}�O���q���E�����͎���̒�ł�����ł��̂ꂽ�B���ꂪ��B�ł͐�Ό����Ȃ��M�t�`���E��L�x���^
�e�n�ɉ����Ė߂��Ă���̂�����A����͂����~�߂��Ȃ��B���w�𑲋Ƃ��邱��̕W�{���ɂ́A���{�̓y����Ƃ���Ă����`���E200��̂W��
�ȏオ���Ԃ悤�ɂȂ��Ă����B
���ꂩ����I�A���̎��͋�B����͐�`�����ꂽ���l�s�ɋ����\���Ă���B10
�N�قǑO����A���Ƀi�K�T�L�A�Q�n��c�}�O���q���E�������p�������n
�߁A�嗤����̐l�דI�A�����Z���ȃA�J�{�V�S�}�_���ƕ���ŁA�����Ƃ����ԂɌ�������p�x�̍ł������`���E�ɂȂ����B����A���̎���̃`���E���͋�B��
��炵�����N���セ�̂��́A�����ɏZ�݂�������ɂ͑z�������Ȃ��������i�ł���B���\�N�������Ă͂��ƖK�˂Ă��Ă��ꂽ���J�m�Ƃ����C�����Ȃ���
���Ȃ����A��͂��a���̕����͂邩�ɋ����B�W�{�̕��X�����Ƃ��������������܂ő����Ă����Ƃ��Ă��A���݂̋�B�̎q�����ɂ͎��̂悤�ȘB���p���g���Ȃ�
�Ȃ��Ă��邱�Ƃ́A���Ȃ��Ƃ��m���ł���B
�`���E�Ɍ��炸���܂��܂ȍ����̖k�オ�b��ɂ̂ڂ�B�����ɂ��Ă͂�
���c�_���c��ɂ��Ă��A���{�̉��g���X���͂��͂⓮����������̂悤�Ɏ��ɂ͌�����B�i�K�T�L�A�Q�n�̗c�����H�ׂ�~�J����J���^�`�A�c�}�O���q��
�E�����̗c�����D�ރp���W�[�́A�֓��n���̎s�X�n�ɂ�������ł�����B�ނ�̕��z�g���j��ł������̂��~�̊����������̂ł���A�C�����オ��Ζk�i��
��͓̂��R�̂Ȃ�䂫���Ƃ������Ƃł���B
�ꌩ�A���͂̎��R�����₩�ɂȂ����悤�ɂ݂��邪�A���Ԃ��ł͂�
���B����n�̐��������ꂾ���̑��x�Ői�o���Ă���Ƃ������Ƃ́A�k���n�̐����͓��������̃y�[�X�Ō�ނ��Ă���Ƃ������Ƃ��B�L�����́A�����P�ɁA�V����
����悤�ɂȂ������͖̂ڂɂ��₷�����A���Ȃ��Ȃ������̂͋C�Â��ɂ����Ƃ����l�ԐS���̗��Ƃ����ɉ߂��Ȃ��B�����A�ߋ��̑����̐�ł��A�قƂ�ǒN��
���C�Â���Ȃ��܂܋N���Ă����B���Ӑ[���Ď����Ă��Ȃ��ƁA�Ƃ�ł��Ȃ����ԂɂȂ��Ă��܂����˂Ȃ��\��������B
�����Ƃ��A�P���Ȑ}���ɂ͓��Ă͂܂�Ȃ��A���g���̔����Ƃ��v���錻��
������������B���Ƃ��A�E�X�o�V���`���E�̐��͊g��B�{���͊���n�̃`���E�ł���͂����A�C���㏸�ƂƂ��ɕ��z���ɍL������A��n�ɉ��肽��A����
�`���E�Ƃ͋t�s���Ă���B�C��ω��ɂ����̂ł͂Ȃ��A�c���̐H�������T�L�P�}����������r�n���L���������Ƃ��������Ƃ�������L�͎�����Ă���悤��
���A�L���͈͂ɓ����悤�ȌX�������邱�Ƃ��炷��ƁA���ꂾ���������Ƃ��v���Ȃ��B�k�J�̃`���E�Ƃ��ėL���ȃX�M�^�j�����V�W�~�̕��z�́A�Ȃ�����k�J�n
��ɍL����n�߂Ă���B������́A�{���̐H���Ƃ����g�`�m�L�̔z��ւ��i�o���Ă��āA�H���̓]�����Ă���͖̂��炩���B
���g�����l�ׂ̂����炵�����̂ł���Ȃ�A����͂������čD�܂�������
�Ƃ͂����Ȃ��̂��낤���A������}���Ɏ~�߂�肾�Ă�����l�ނɂȂ����Ƃ��m���ł���B�����̕��z����C�ɏ�����������悤�ȋC��ϓ����킪�g�Ŏ���
�ł���悤�ȋ@�������������Ƃ͎v���Ȃ��BDNA���̂��̂̕ψق�A�G�s�W�F�l�e�B�b�N�ȕω��ȂǕ��q���x���̃`�F���W
�����łɎn�܂��Ă��邩�����ꂸ�A��������������Ɛ����̑��ݍ�p��m�邽�߂̑s��Ȏ����ƂƂ炦�Ă͂ǂ��Ȃ̂��낤�B���ԂƂ��Ă̐����̐i���⑽�l��
�ɔ���`�����X�����������̂��A�Ƃ������z�������Ă����̂��Ǝv���B
|
�����@����
�T�C�G���X���C�^�[
|
2011.6.20
�@No.2 �@
�@�@�u�̏W��������ցv �V�����������N�̈�Ă��� ���@�@�F�@�i
|
���͏��a�Q�W�N��
�܂�ŁA���S�t
��
�����ɂ͍��x�o�ϐ������n�܂��Ă���A���s�s���̏��w�Z���ӂɂ������n�A���c�A�т��N�X�Z��n�ɕς���čs���̂�ڂ̓�����ɂ����B����ł����w����
�ċx�݂͖����U�����璎���ŁA���t���O�̐��ⓔ�ɑO������J�u�g���V��N�����������ɍs�������������B���w�Z�̂T�A�U�N�̓`���E�ɋ������o�āA
���s�s���̃`���E��S��ޏW�߂Ă�낤�Ə��w���Ȃ�Ɏv���Ă����B���������w���ɂȂ�Ə�ɂ͏オ���邱�Ƃ�m��A�ނ̕W�{���ɖ�O�Ō������Ƃ��Ȃ��E�X
�o�V���`���E�������Ă���̂����ċ������B�e��Z�ɍ̏W�ɘA��Ă����Ă��炤�̂͂��邢����Ǝv�����B���w�Z�Ɠ����~�n�ɂ��������u�Б�w�̍����W�{�W
�����w�Ղ̍ۂɌ��ɍs���A�q�T�}�c�~�h���V�W�~�̕W�{�������Ă�������B�������ʂ�V�T�C���͂��̎����߂ċ����
���B�قǂȂ��q�T�}�c�~�h���V�W�~�̐����j���𖾂��ꂽ�Ƃ̋L�����V���Ɍf�ڂ���A���̃`���E������Ȃɒ��������̂ŁA����ɂ��ꂪ���s�ɂ��邱�Ƃ�m��
�ċ������B���s�k�R�̐������q�T�}�c�~�h���V�W�~�̐��n���Ƃ́A���w���̎��͒m��R���Ȃ������B�����T�`�P�O�Ώ�̐���͂���܂Ŗ��m���������{��
�`���E�̐��Ԃ����X�Ɖ𖾂��ꂽ����������߂����ꂽ�Ǝv���B��ʂ��s�ւŖ��J��ȏꏊ�����{�ɂ�������c���Ă���A�����ł̍̏W�Ɗώ@�ɂ���ē���ꂽ
�V�����m���͎R�̂悤�ɂ������B
�@��
�݂͂ǂ����낤�B�������N�͂�
�Ȃ��Ȃ��ċv�������A��w�̗��w�������Ȋw�Ȃɍ������D���œ��w����w���ȂLj�l�����Ȃ��B�g�̉��̍����͋ɒ[�ɏ��Ȃ��Ȃ苻���̑Ώۂ��̂��̂��g�߂�
���Ȃ��Ȃ����B�n�����D��┎���ق������̏W����J�Â��Ă��A�ȒP�ɂ͐V���Ȓm���������Ȃ��Ȃ����������̏W����Ӌ`�����̊y�������q���B�ɓ`���邱��
���ł��Ȃ��B��������̑�l�����́A�����������y���v���o�����̂܂܈��������āA�����悤�Ɏq���B�������������Ƃ��Ă���B�q���B���g�̉��̍�����
�̏W���ċ^��Ɏv�����Ƃ̂قƂ�ǂ͂��łɑ�l�B�����炩�ɂ��Ă��܂����B����ł͍������N�͈炽�Ȃ��B�������N���痝�ȍD�������܂�A�Ȋw�҂����܂��
�Ƃ������͂������݂��Ȃ��悤�Ɏv����B
�@��
�͉�������悢���낤���B��
�����̏W���`�Ԃ̍ו��ׂĕ��ނ�����A�n��ɂ���Ă킸���ɈقȂ����̈Ⴂ�ׂ���A���ǂ��ɉ��������������ׂ��肷��悤�ȁA�]�O�̍���
�����͖ʔ����Ȃ��̂ł���B���͎������Ǝv���B�q���͗��Ȏ�������D���ł���B��w�̌��J�u���̎��ɂ͏����ȊȒP�Ȏ����ł��q���B�̎Q����]�҂��E�����A
�L���L���P���ڂ����邱�Ƃ��ł���B���ݓI�ɂ͍����D���̒j�̎q�͂�������̂ŁA��l�����̂悤�Ȏw�������Ă݂Ă͂ǂ����낤�B��O�̑̌��ŕs�v�c��
�Ɗ������������ɏ������߂Ă�������B�s�v�c�Ȏ����̐����闝�R��J�j�Y���ɂ��ĉ����𗧂Ă�����B���������������Ƃ������ɂ͂ǂ̂悤�Ȏ�����
�����炢�����l��������B���D��┎���ق͎q���B�Ɏ���Ŏ��������A���ɂP�A�Q������̎w�����s�Ȃ��B
�@��
���ȑ��u��ޗ����g�킸�A����
�łł�������͂�������B�ŋ߂��̂��Ƃ����g�Ŏ����ꂽ�͕̂���s���搶�ł���B�u�A�Q�n�`���E�̗c���͕\�ʂ��U���U�������ꏊ��匉��������ɊD�F��
匂ƂȂ�v�Ƃ��������𗧂āA���������Ō����ɏؖ����ꂽ�B������_���I�ɑg���ĂĂ����v���Z�X�����炵���B������ӂɂ���ޗ��������g���ē���ꂽ
�������ʂ��ꗬ�̍��ێG���Ɍf�ڂ��ꂽ�B�������Ƃ��c�ɍs�����ɑ�w�̃L�����p�X����������ŁA�s�v�c�Ɏv�������ɂ������o���킷�B���ꂩ�痧�Ă���
���ɂ́A�Ⴆ�Ύ��̂悤�Ȃ��̂�����B�u�n���r���J�}�L���̗c���͉��F���Ԃ�I��ŏW�܂�v�u�����V���`���E�̖{���̐����n�͍r��n�ł���v�u�S�}�_��
�`���E�̗c��������z�~����͖̂̍����ɗ��t���Ȃ����ł���v�B�������ؖ����邽�߂̎����͏����l����Αg�ݗ��Ă邱�Ƃ��ł���B���w�������l�܂�
�P�l�Ŏ����Ɏ��g�ނ��Ƃ��ł���B���Ƃ͂����ɃG���K���g�Ɋm���ɏؖ����Ă݂��邩���r�̌����ǂ���ł���B
�@��
����PCR���u��DNA�V�[�P���T�[���g�����Ƃ��D�ꂽ�����ł͂Ȃ��B���
�K�v�Ȃ͎̂��R��s
�v�c�Ɏv��������{�����ƁA���ɉ������ؖ����邽�߂̎����̑g���Ă��l�����邱�Ƃł���B�V�����������N�́u�̏W��������ցv�A���ꂪ���ȍD������āA
�n���͖L���ȉȊw�҂���邱�ƂɂȂ�̂ł͂Ȃ��낤���B�B��̊뜜�́A�̏W�ƕ��ނ������Ă�����l�ɂ��ꂪ�{���Ɏw���ł��邩�Ƃ����_�ł���B
|
���@�F�i
���{����w��w�@���w�n�����ȁ@����
|
2011.5.8
�@No.1 �@
�@�@��
�������҂͂Ȃ��������Ă��邩�H ��@�V�@�ȁ@�O
|
�@���̂Ƃ���A���D���̎�҂�����
���Ȃ��Ȃ��Ȃ��Ă����A�Ƃ����Q�������X�ŕ������B���{�̎R�삪�J�����b�V���ŁA���̂��݂��������������Ƃ����낤���A���ȊO�̒ᑭ�H�ȃQ�[���̔�
���Ȃǂ���҂̋��������Ȃǂ����Ȃ��Ȃ����̂�����Ƃ��Ă������悤�B
�@�������A���͂���ȊO�̂����Ƒ�
���Ȍ����́A������䂪���̍����w��majority���A�ÐF���R�Ƃ����L�ڕ��ފw�Ő�
�߂��Ă��邱�Ƃɂ��A�Ǝv���Ă���B�Ⴆ�A�Ԗ�����ߎ���́u����͐ԁv�Ə������A�ʔ������Ƃɂ́A����̎�̐Ԃ͐F�f�n�A�����͍\���F�ł����
�������邵�A�������i�܂��͐ԁj�F�ł��F�f���قȂ�ȂǁA�P�ɋL�ڂ�ǂ����ł͕�����Ȃ��B���̂悤�ȗ�͑��ɂ��R�قǂ���A�ŏI�I�ɂ͂����̐F��
�x�z�����`�q�̓���Ƃ��̔����̐���@�\��m��K�v�����邱�Ƃ͂����܂ł��Ȃ��B���̂悤�Ȏ����ɁA�����̑��l���̈�[���B����Ă���A�i���̋@�\�̉�
���ɂ��v������ł��낤�B���₱���̉�͉͂\�ƂȂ����B
�@���ꂾ�������w�������̐i������
���A���Z�Ȃǂ̋���̏�ł��A���Ȃ荂�x�ȐV���������w�������Ă���̂�����A��قǂ̗�O�������A�����A���݂̓��{�́g�������w�h�̌����@�ւɓ�����
�ꐶ�����������Ƃ����҂��������A����ɔ����A�A�}�`���A�̒����̖ړI�ӎ��ɂ��ω��������͓̂��R�̋A���ł���B�^�S�����̍�N�x�̍b���̉�ڂ̍���
�݂�ƁA��N�L�ڂ��ꂽ�V��̃��X�g����̂ŁA����DNA������Ŏ��X�Ƃ������g�{���ɍ�
���͖ʔ����h�@�Ǝ����v�������͂قƂ�Ǐo�Ă�
�Ȃ��B
�@���́A���ފw�͕K�v�s���Ȃ���
���Ǝv���Ă͂��邪�A�����܂Ői�����������w�S�̂���݂�ƁA����܂ł̕��ފw��10%���炢����A�����w�S�́i������
��ʓI�ɂ����A�����w�j����݂ď\���ł���Ǝv���B����ɂ́A�����ق��w�̍����������̒�������Ȃ��Ƃ����{�ɑ����A���ފw�҂����Ń|�X�g��Ɛ肵��
�����Ƃ��K�v�ł���B��������Β��D���̎�҂̑����́A�����ɂ͂����ƁA�����Ɩʔ������Ƃ��R�قǂ���̂�����A���낢��ȐV�����Z�p��l������g�ɂ�
���A�V���������w�̓��i�݂����Ɗ�����̂ł͂Ȃ����Ǝv���B�����w�����������i60�N���́j�A�̍]��s��v�搶���u��
�吨�̂ЂƂ�����Ă��邱�Ƃ͂��ȁB�����Ŗʔ������Ƃ��݂��A�����ʔ�������v�Ƃ���ꂽ���A���⍩���w�ɂƂ��āA���ꂪ�d�v�Ȃ̂ł���B�v����
�ɁA�l�܂˂͂���ȁB�����œ����J���A�Ƃ������Ƃł���B
�@�����̓r�����Ȃ����l������݂�
�A������Ȃ����Ƃ��炯�ł���B�����͑S������majority���߂�̂�����A���������ł͐i
���̖{����������Ȃ��Ǝv���B���̂Ƃ���A���f���������g���āA�V�������ۂ�A����܂őz�����ł��Ȃ������悤�ȋZ�p�������Əo�Ă��Ă���B�����^
������A�Ƃ����̂ł͂Ȃ��B�Ⴂ����̕��͂��������܂����p���A���ǂ��A�X�ɂ͎��g�œƎ��Ȃ����������āA�ʔ������̐��E�����ۂƂ��Č��邾���ł�
���A�ǂ̂悤�Ȏd�g�݂ł��̂悤�Ȃ��Ƃ��N���Ă���̂��A����������A���ۂ̖{�����𖾂��Ăق����B�����Ȃ��Ďn�߂č����w�͐����Ȋw�����ԂƔF�߂��
��̂ł���B���̂悤�Ȋϓ_����݂āA����DNA������̖����͂��ꂩ��܂��܂���
�傫���Ȃ�ł��낤�B
|
���V�@�ȎO
������w�E�L����w���_����
|
|
|
|
|













